Az elmúlt kétszáz év felemelően szép eredményeinek dacára egészen az 1980-as évekig néhány gyümölcsöző rövid időszakot leszámítva az evolúciós biológia és a fejlődésbiológia szinte teljes elszigeteltségben fejlődött, más célokkal, más metodológiával. Az utóbbi kicsit több mint három évtized hozta meg az áttörést a modellorganizmusok morfogenezisének a „klasszikus” módszerek mellett jórészt új, molekuláris genetikai módszertannal történő megismerése, a molekuláris technikák egyre bővülő tárháza egyre inkább integrálódott az evolúciós kutatás különböző ágaival, létrehozva az interdiszciplinaritás egyik csúcsát jelentő evolúciós fejlődésbiológiát (evolutionary developmantal biology= EVO-DEVO).
A homeotikus mutánsok már régóta foglalkoztatták a kutatókat. Bateson: állati variánsokra hívta fel a figyelmet az 1890-es években, ahol egyik részt egy másik helyettesít - „szörnyek” („monsters”) névvel illette őket, ő alkotta meg a homeotikus jelzőt is.
Bridges 1915-ben fedezte fel az első ilyen mutánst Drosophilában, ez volt a bithorax mutáció. Már a korai tanulmányokból is érzékelhető volt, hogy SEJTSORSOK (CELL FATE) KÖZÖTT VÁLASZTANAK, TEHÁT SZELEKTOR GÉNEK.
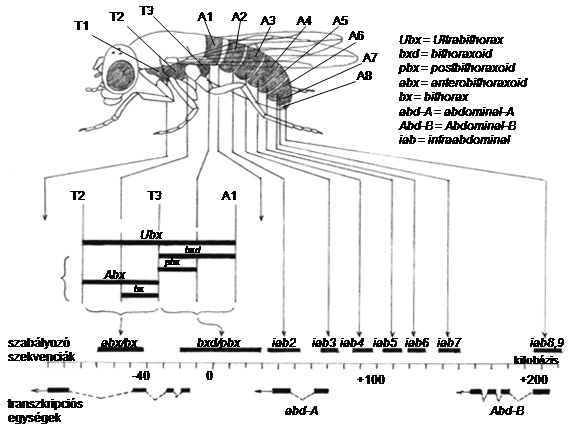
Kétségtelenül döntő jelentőségű volt az állati homeobox-gének Hox-klasztereinek felfedezése és jellemzése az 1970-es és ’80-as években. A Hox-gének a homeotikus szelektorgének legismertebb osztálya, amely szabályozza a különféle testrészek mintázatképzését. Drosophilában a Hox-gének két gén-komplexben (gén-klaszterben) helyezkednek el a 3. kromoszómán, elnevezésük: bithorax-komplex (BX-C) és Antennapedia-komplex (ANT-C), de szokás a két komplexet összefoglaló néven HOM-C-ként említeni (más fajokban, mint a lisztbogárban, valóban egy, összefüggő komplex van). A BX-C úttörő genetikai analízisét E. B. Lewis végezte el, több mint három évtizedig tartott, az ANT-C analízise a’70-80-as években folyt, Th. Kaufman vezetésével 12. ábra.
10.12. ábra - Lewis modellje a bithorax-komplex (BX-C) szabályozására. A második tori szegment (T2) az alapállapot, minden utána következő szegmentben újabb gén aktiválódik. Az utolsó szelvényben minden gén aktív
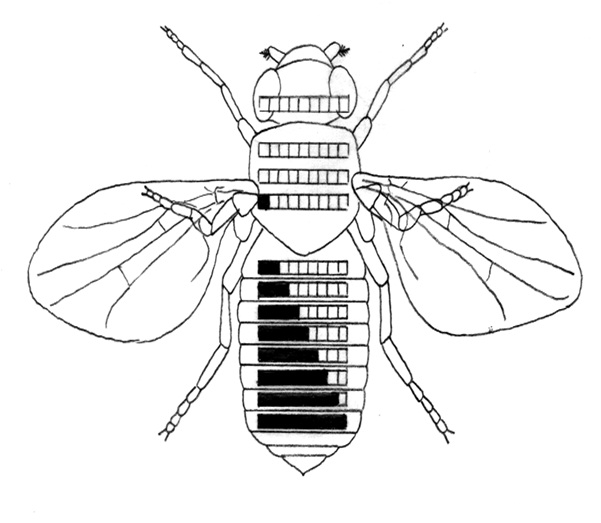 |
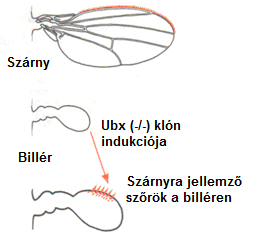
A kutatók azt találták, hogy a két komplex génjei a szelvények egyediségét szabályozzák az A/P tengely mentén, helyzetük pedig megfelel kifejeződésük rendjének: a kromoszómán ugyanaz a génsorrend, mint a kifejeződési sorrend a testben. A Hox-gének szabályozzák a szelvények egyediségét metazoákban az elülső-hátsó (A/P= anterior-posterior) testtengely mentén. A Hox-gének mutációi gyakorta okozzák egyik szelvénytípus transzformációját egy másik típussá. Drosophila melanogasterben például, az Ultrabithorax (Ubx) gén mutációja a harmadik torszelvényt második torszelvénnyé transzformálja. A transzformáció nyomon követhető: a harmadik torszelvény billérje szárnnyá transzformálódik 13. ábra.
Egy másik Hox-gén, az Antennapedia (Antp) mutációja hatására az Antp-fehérje a lábak mellett azokban a sejtekben is kifejeződik, amelyekből csáp (antenna) fejlődne, így a csápok helyén lábak lesznek. Az Antp-gén általában csak a második torszelvényben (T2) fejeződik ki, ahol a T2-re jellemző képletek, köztük lábak kialakításában vesz részt (A transzformációkkal kapcsolatosan elmondottak némi egyszerűsítést jelentenek, de az egyszerűsítések a lényeget nem érintik.) 14. ábra. Lásd a 15. ábra és a 16. ábra mutánsait is!

Arra is fény derült, hogy a komplexek nyolc génje közeli rokon: ugyanazon géncsalád tagjai és a kódolt fehérjékben megvan egy bizonyos aminósav-szekvencia, amely a DNS-hez köt; későbbi elnevezése homeobox (a gén 180 bázispárnyi szekvenciája) ill. homeodomén (a DNS-kötő fehérjedomén). A felfedezések támogatták Lewis korábbi, ’60-as évekbeli javaslatát: a Hox gének más géneket szabályoznak.
10.15. ábra - A bithorax-komplex szabályozó szakaszainak mutációi. A kifejlett egyed szelvényekből, a szelvényeken belül elülső és hátsó kompartmentekből áll. A kijelölt szakaszok a szabályozó (=regulátor) régiókat, azt alattuk lévő folyamatos vonal a komplex 300 000 bázispárnyi hosszát jelöli. Alul azt a három transzkripciós egységet jelöltük, amely a komplex három homeotikus fehérjéjét kódolja (a klasszikus „ kódoló” szakaszok a komplexnek kisebb részét teszik ki, a nyilak az átírás irányát mutatják)
 |
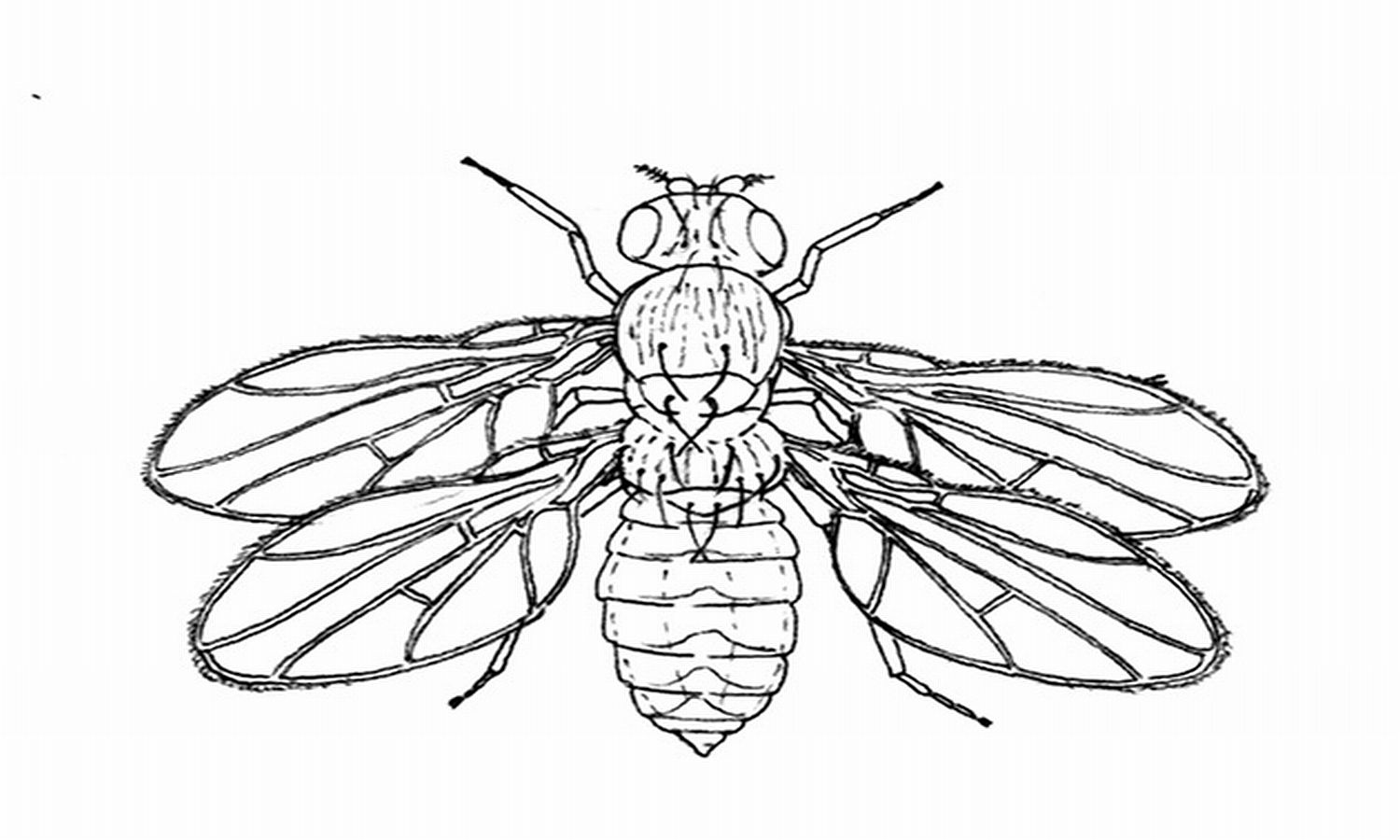
10.16. ábra - A híres négyszárnyú légy, amelynek három független mutációja a 3. torszelvénynek teljeskörű 2.-ká transzformálódását eredményezi
 |
Más kutatók hasonlóan jelentős felfedezést tettek: Minden más állati törzsben jelen van Hox-gének egy készlete. A vizsgált újabb törzsek Hox-génjeinek is van a Drosophila génekéhez nagyon hasonló homeoboxa (homeodoménja), sőt, a gének sorrendje és orientációja is a legtöbbször hasonló (bár a legtöbbször a génkomplex egy, folyamatos egységet alkot).
Az emlősöknek négy Hox-gén komplexe van (elnevezésük Hoxa, Hoxb, Hoxc, Hoxd), és a Drosophila melanogaster 8 génes komplexével szemben az emlős Hox-komplexek 13 különböző Hox-gént tartalmaznak (bár nincsen meg minden gén minden komplexben).
A Hox-fehérjék vagy Hox-mRNS-ek kifejeződését kimutató festések kimutatták, hogy a Hox-gének a A/P kifejeződési mintázata megfelel a mutáns fenotípusoknak: az Ubx-gén a T3-, és elülső potrohban fejeződik ki, mint az tudott volt a mutánsok analíziséből. A gerincesek Hox-gén kifejeződése, bár komplexebbek, szintén specifikus A/P mintázatot mutat. Külön fejezetben foglalkozunk majd a Hox-gének evolúciójával, megemlítjük, hogy a Bilateriákkal, a komplex állatokkal testvércsoportot alkotó Cnidariák (medúzák, korallok) esetében is megtaláltak kis számú, Hox-hoz hasonló gént. Jónéhány új Hox-gén jelent meg a Bilateraliákhoz vezető vonalon, amelyek új Hox-osztályokba sorolhatók (homeodomén struktúrájukat figyelembe véve), így minden bizonnyal lehetővé vált az elülső-hátulsó tengely finomabb differenciálódása.
A Hox-génekkel kapcsolatos felfedezéseket nem lehet túlértékelni az állati diverzitás evolúciójának megismerése szempontjából. Ez volt az első olyan alkalom, hogy egy közös fejlődésgenetikai keretbe lehetett foglalni minden metazoa ontogenezisét és evolúcióját. Ezeket a felfedezéseket megelőzően nem volt olyan kutató, akiben akár felmerült volna, hogy a gerincesek és gerinctelenek ennyire mély közös fejlődésgenetikai alapokkal rendelkeznek. Ezek a felfedezések ráadásul erjedést indítottak meg: elindították a közös alapok keresését a fejlődés minden elképzelhető vonatkozásában: a hát-hasi mintázatképzéstől a végtagok és szemek kialakulásáig. A morfológiai evolúció tanulmányozása sajátos módon szinte felfoghatatlan mértékű konzervativitást tárt fel, nem változatosságot, ami az evolúció lényegi sajátsága. Azonnal felmerült a kérdés: Mi a testtervek különbözőségének alapja? Hogyan játszhatnak minden jel szerint konzervatív genetikai faktorok fontos szerepet ezekben a különbségekben?
A kérdésekre a későbbiekben hosszabb választ adunk, most csak dióhéjban válaszolunk:
A szelvényszintű Hox-génkifejeződés különbségei látványosan korrelálnak az állati testterv evolúciójával. Doménjaik eltolódásának drámai hatása lehet a morfológiára. Kifejeződési módjuk az egész állatvilágban hasonló, azaz a kromoszómán való elhelyezkedésük kolineáris kifejeződésük sorrendjével a testtengely mentén (zootípus).
A Hox-gének transzkripciós faktorokat, vagyis olyan fehérjéket kódolnak, amelyek bizonyos DNS-szabályozó régiókhoz kötnek (más néven cisz-regulátor elemekhez: promóterekhez, enhancerekhez) kötnek. A cisz-regulátor elemek ún. cél (target) gének kifejeződését szabályozzák, amelyek a génhierarchiában a Hox-gének alá vannak rendelve („downstream” gének). A morfológiai változások a Hox-gének által szabályozott gének kifejeződésének változására vezethetők vissza.
A Hox-gének szerepének tanulmányozása az állatok evolúciójában új alapokra helyezte gondolkodásunkat a morfológiai evolúcióról: az új rendszer alapja, hogy egy közös, kisszámú génből álló génkészlet, a „toolkit” génjeinek tér- és időbeli változása a morfológiai evolúció elsődleges oka.