Elsősorban Ed Lewis-nek az ötvenes években (ld. Hox-gének, következő fejezet) az Egyesült Államokban, valamint Christiane Nüsslein-Vollhard és Erich Wieschaus-nak a hetvenes évek végén Németországban megkezdett kísérletei nyomán indultak meg azok a kutatások, amelyek azokra a génekre irányultak, amelyek a Drosophila melanogaster testtervének kialakításában részt vesznek, és a pozícionális értékek kialakításának letéteményesei 10. ábra. A legtöbbjüket azon az alapon találták meg, hogy fontos szerepet játszottak az embriók és a felnőtt állatok szegmentális mintázatának kialakításában. Ezekből a kutatásokból sarjadt ki a modern molekuláris genetikának az a tudománytörténeti jelentőségűnek bátran nevezhető forradalma, hogy az állatvilág jelentős része ugyanazt a nagyon konzervált molekuláris rendszert használja a pozícionális értékek „jegyzésére” a sejtekben a test hosszanti tengelye mentén, és a gének közül néhány a gerincesek végtagjainak kialakításában is részt vesz. A rendszerről a későbbiekben dióhéjban lesz szó, az érdeklődők a részleteket fejlődésbiológia könyvekben megtalálják.

A fejlődésgenetikai analízisének az a lényege, hogy bizonyos mutánsok fejlődésének megváltozásából következtetünk annak a génnek a normális funkciójára, amelyben a mutáció történt.
Az A/P tengely kialakulását szabályozó hierarchia és főbb sajátságai Drosophilában:
-
Anyai transzkripciós faktorok grádiense alakul ki
-
A gap gének transzkripcionálisan aktiválódnak és keresztbe szabályozzák egymást
-
Az egyes pair-rule sávok kialakulását az anyai és gap fehérjék szabályozzák
-
A pair-rule és szegmentpolaritási gének pontos kifejeződését más pair-rule és szegmentpolaritási gének szabályozzák 11. ábra
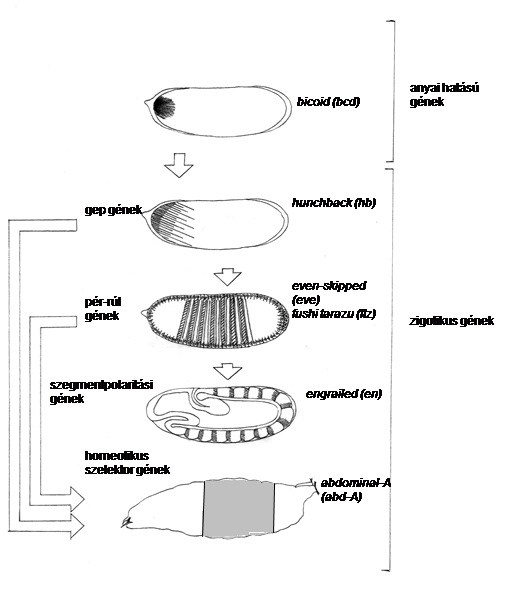
10.11. ábra - Különféle génkészletek egymásra következő kifejeződése alakítja ki az A/P tengelyt. A megtermékenyítés után az anyai géntermékek transzlálódnak, és pozicionális információt biztosítva aktiválják a zigotikus géneket. A gep (gap=rés) gének regionális különbségeket definiálnak, amelyek a pair-rule (=pár szabályozó) gének periodikus mintázatává transzformálódnak. A pair-rule gének definiálják a paraszegmenteket. A szegmentpolaritási gének munkálják ki a szelvények mintázatát, majd a homeotikus szelektor gének határozzák meg a szelvények egyediségét
Általános tanulságok a szegmentációs gének tanulmányozásából
-
1. grádiensekre adott a koncentráció-függő válasz a különböző szintű és a küszöbérték által kiváltott válasz egyaránt előfordul a gap-gének és bicoid fehérje esetében
-
2. aktivátorok és represszorok egyaránt meghatároz(hat)ják a gének kifejeződési mintázatát- a térbeli represszió mindig fínomabb mintázatokat „vés ki” nagyobb doménekből
-
3. sok gént két vagy több aktivátor vagy represszor regulál. A kombinatorikus szabályozás nagyobb specificitást enged meg, és diverzebb térbeli mintázatok alakulhatnak ki
-
4. több független regulátorelem szabályozza a géneket. Az egyedi elemek egyedi doméneket szabályoznak
-
5. a gének szekvenciális aktiválása a szabályozó hierarchiában tökéletes időzítést tesz lehetővé
Az elsődleges és másodlagos mezők génkifejeződésének logikája
A fejlődést szabályozó mechanizmusok maguk is evolúciós termékek, így tükrözik az őket létrehozó evolúciós folyamatokat.
A regulációs hierarchiák architektúrájára, a génexpresszió mechanizmusaira és a celluláris mezők felosztására, valamint a toolkit gének szabályozó szerepére három fontos szabályszerűség állapítható meg:
1. A növekvő embrió fejlődése térben és időben rendezetten, sorban megjelenő koordinátarendszereken múlik
-
=> bármely felnőtt struktúrát hierarchiák egy sorozata determinál, amely az embriót egyre finomabb részekre osztja. A másodlagos mezők helyét A/P (Hox) <=> D/V majd önálló koordinátarendszerük határozza meg, ezen belül lokális génkifejeződési doménjük alakul ki
-
A FOLYAMATOK TÉR- ÉS IDŐBELI SZEGREGÁCIÓJA => MODULARITÁS, INDIVIDUALIZÁCIÓ
2. Az egyes gének cisz-regulátor elemeinek modularitása lehetővé teszi a génkifejeződés tér- és időbeli finomszabályozását => különböző kontextusokban specifikus funkciója lehet az egyes géneknek
-
Ugyanazt a jelátviteli útvonalat pl. A jelfehérje nagyon különböző szövetekben indíthatja be
-
A fehérjék funkciója kontextusfüggésük miatt nem írható le csupán a fehérje alapján
-
MINDEN EGYES SZABÁLYOZÓELEM EGY FUNKCIÓ
-
MODULARITÁS: 1. tükrözi a génkifejeződés fejlődését 2. egyik funkció változhat, míg a másik megmarad => génfunkciók disszociálhatnak, új funkciók alakulhatnak ki
3. A térbeli mintázatok kombinatorikus szabályozás termékei
-
Pl. Végtag helye
-
A KOMBINATORIKUS SZABÁLYOZÁS KULCS A SPECIFICITÁSHOZ ÉS A DIVERZITÁSHOZ. PL. LEHETŐVÉ TESZI, HOGY DISZKRÉT SEJTPOPULÁCIÓKBAN FEJEZŐDHETNEK KI GÉNEK.
-
A DIVERZITÁS VÉGTELENNÉ VÁLIK A KOMBINÁCIÓK RÉVÉN
A fentiekhez a lehető legszervesebben kapcsolódik a regulatory evolution eszméje, amelyről a következő fejezetben lesz részletesen szó, elsősorban a szekvencia oldaláról megközelítve a most tárgyalt elveket.