Az állati genom gének tízezreiből áll. A „housekeeping” – háztartási - gének minden sejt számára esszenciális funkciókat kódolnak
Vannak specializált sejtek vagy szövetek génjeinek funkciójáért felelős gének (immunsejtekben pl.), analógiaként „a ház szobáiban dolgoznak” kifejezést alkalmazhatjuk.
MINKET OLYAN GÉNEK ÉRDEKELNEK, AMELYEK A „HÁZ” FELÉPÍTÉSÉT IRÁNYÍTJÁK - A „TOOLKIT” („SZERSZÁMOSLÁDA”) GÉNJEI, AMELYEK MEGHATÁROZZÁK A TESTTERVET, ÉS A TESTRÉSZEK IDENTITÁSÁT, MINTÁZATÁT, ELHELYEZKEDÉSÉT ÉS SZÁMÁT.
Mint láthattuk, két alapvető forrás volt a „toolkit” génjeinek azonosításához:
-
- „szörnyetegek”- ritka, spontán felbukkanó mutánsok
-
- szisztematikus szkrínek mutagénekkel
Felosztás: Hol és hogyan funkcionálnak a „toolkit” gének?
-
i. a mezők egyediségét (pl. szelvények, függelékek)
-
ii. a mezők normális kialakulását, kompartmentalizációt (pl. szervek és függelékek)
-
iii. sejttípusok kialakulását (izom, neurális sejtek)
-
iv. a testtengelyek kialakulását (ld. előbb)
szabályozzák.
(i) Mező-specifikus szelektor gének
Egész struktúrák kialakulását és/vagy mintázatképzését szabályozzák. Ektopikus kifejeződésük esetén ektopikus szervek. Az eyeless gén funkciója ha hiányzik - szem nélküli légy. A gén molekuláris elemzése alapján - homeobox-gén, a Pax6 rokona 17. ábra.
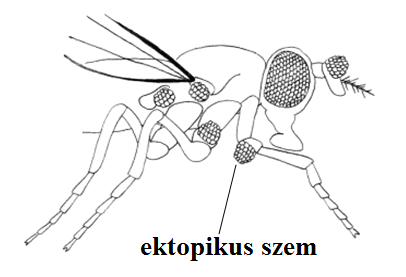
További mezőspecifikus szelektorgének: a Distal-less (Dll), amely a lábak, szájszervek, antennák, genitáliák proximo-disztális fejlődéséért felelős. Hiányában mindezen képletek disztális elemei hiányoznak. Homeoboxot tartalmaz.
A vestigial (vg) és scalloped (sd) együtt hat, hiányukban a légynek nincsenek szárnyai és haltérái. Termékeik DNS-kötő fehérjekomplexet alkotnak. A tinman gén hiányában nem fejlődik szív- különleges homeobox-fehérje.
(ii) Kompartment szelektor gének
A kompartmentalizáció olyan határok létét jelenti az élőlény fejlődési mezőin belül, amelyeket bizonyos sejtek utódjai (klónjai) sohasem lépnek át. A határok a legtöbb esetben nem köthetők anatómiai értelemben vett határvonalakhoz.
Az engrailed minden szelvény hátulsó kompartmentjében hat, folyamatosan az élet során, így a szelvényből származó képletek hátsó kompartmentjében is kifejeződik. A szárnyban és az embrióban „poszteriorális” identitást határoz meg. Mutánsaiban az anteriorális (elülső) rész tükörszimmetrikusan jelenik meg. Homeodomén-transzkripciós faktort kódol. Az apteorus gén terméke a szárnydiszkuszt osztja fel háti és hasi kompartmentekre.
(iii) Sejttípus-specifikus szelektor gének
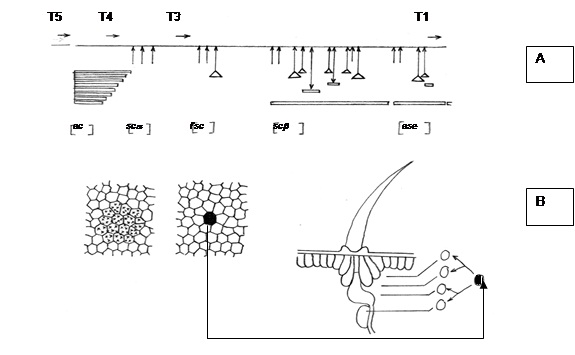
Proneurális gének kifejeződése sejtklaszterekben történik, amelyekből egyetlen prekurzor szelektálódik. Az achete-scute komplex kódolta transzkripciós faktorok homo- és heterodimereket alkotnak egymással és más hasonló jellegű fehérjékhez és célgénekhez kötődnek, iniciálva a neurális sejtsors kialakulását 18. ábra.
10.18. ábra - Az achete-scute komplex (AS-C) térképe. A nyilak azonosított mutációk helyét jelzik. Az üres fekvő oszlopok deléciókat jelölnek, a háromszögek inszerciós transzpozonmutánsokat. A T5, T4, T3, T1A átírt régiókat jelölnek. A nyilak az átírás irányát jelölik. A genetikai lókuszok nevét a zárójelben tüntettük fel: ac: achete, sc-a, sc-b: scute, l'sc: lethal of scute, ase: asense. B: A scute T4 gén kifejeződésének korlátozódása az epidermiszben egy sejtcsoportról egyetlen sejtre, amelynek osztódása 4 sejthez vezet, melyek egyike sem fejez ki T4-fehérjét
 |
(iv) Testtengelyek kialakulását szabályozó gének
Az embriológusok régóta keresték az állati testszerveződés alapelveit
Főbb területeik:
testtengelyek kialakulása
csíralemezek kialakulása
függelékek, szelvények polaritása
Elsődlegesen manipulatív és ablációs technikákkal: diszkrét szervező régiók, organizátorok
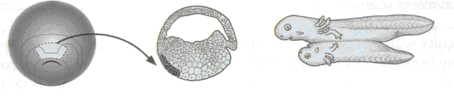
Kimutatták, hogy a korai kétéltű gasztrulában a blasztopórus háti ajka az egyetlen önszervező, önálló differenciációra képes régió. Az organizátor (kétéltűeknél) tehát definiálható olyan szövetdarabként, amelyet egy idegen embrió hasi oldalába ültetve másodlagos háti testtengelyt és fejet indukál. Spemanni alapokon állva, ma a Spemann-organizátornak három alapvető sajátságát emelhetjük ki:
Az organizátor utódsejtjei alakítják ki az embrió axiális (tengely) struktúráit.
Az organizátor sejtek más sejteket toboroznak, amelyek részt vesznek axiális és paraxiális struktúrák kialakításában.
Neurális szövetet indukálnak.
Az organizátor sejtjei további fontos szerepet játszanak a gasztruláció mechanikájában 19. ábra
Drosophilánál megismert anyai és zigotikus, a korai testszerveződést befolyásoló gének, homeotikus gének.
10.19. ábra - Az organizátor régió egymástól távol eső fajokban messzemenő megegyezéseket mutat strukturálisan, funkcionálisan és molekuláris szinten is
 |
Intenzív szkrínek (screen) az embrionális és felnőtt testszerveződést befolyásoló gének azonosítására, és fehérjéik strukturális, funkcionális és expressziós analízise a genetikai toolkit több általános sajátságát tárta fel:
-
1. Az összes gén kis részét alkotják - kevés gén hat specifikusan a fejlődésre
-
2. Legtöbb toolkit gén vagy transzkripciós faktort, vagy jelátviteli útvonal komponenst kódol - a toolkit gének általában más gének kifejeződésére hatnak
-
3. A toolkit gének idő- és térbeli kifejeződése gyakran egybevág azzal a régióval, amelyben a gén funkcionál
-
4. A toolkit gének osztályozhatók a fenotípusok alapján, amelyeket mutációjuk okoz - hasonló mutáns fenotípusok gyakorta arra utalnak, hogy a két gén ugyanazon az útvonalon hat.
-
5. Sok toolkit gén rendkívül konzervatív, különböző törzsekben is megvannak homológjai
Evolúciósan konzervált fejlődési útvonalak
-
A toolkit gének termékei: transzkripciós faktorok és jelátviteli útvonalak fehérjéi. A JELÁTVITELI ÚTVONALAK KOMPONENSEI KÜLÖNÖSEN PLEIOTROPIKUSAK
-
A wingless-útvonalnak pl. szerepe van: korai ektodermában, a szegmentek polaritásának kialakításában
-
később, az embrionális ektodermában a láb- és szárny imaginális diszkuszok kialakulásában
-
a lárvális szárnymezőben a D/V (dorzoventális) polaritás kialakításában, a szárny kibontakozásában, és az érzékszervek mintázatkialakításában
-
végül a szem, láb és más szövetek polaritásának kialakításában
AZ ÚTVONAL ÉS KOMPONENSEI MINDENHOL UGYANAZOK, A „KIMENET” („OUTPUT”) MINDENHOL MÁS. Minden fontosabb jelátviteli út komponensnek megvan a gerinces és a Drosophila megfelelője
-
Néhány széles körben elterjedt jelátviteli fehérje fontos szerepet játszik a „klasszikus” organizátorokban
-
A gerincesekből ismert chordin megfelel a Drosophila short gastrulationnak, mindketten a TGF-β család tagjaival lépnek gátló kölcsönhatásba, és mindketten a D/V tengely kialakításában vesznek részt
-
Sonic hedgehog - ZPA helyén fejeződik ki
-
FGF-géncsalád - AER helyén
A morfogenezist szabályozó gének hierarchiákat vagy hálózatokat alkotnak. Ezek a gének a jelátvitelben részt vevő fehérjéket kódolnak, amelyek molekuláris jeleket (ahogy gyakran mondjuk: szignálokat) közvetítenek a sejteken belül és a sejtek között, transzkripciós faktorok, amelyek a jelközvetítő útvonalak hatására növelik vagy csökkentik célgénjeik átírását, valamint strukturális gének, amelyek a fejlődés és élettan folyamatainak tulajdonképpeni kivitelezői (pl. enzimek, citoszkeleton-alkotók). Jónéhány, a főbb szerveket és függelékeket kialakító fejlődési útvonalat szélsőséges mértékben konzervált transzkripciós faktorok szabályoznak. A distalless gén például olyan transzkripciós faktort kódol, amely irányítja a testből kinövő függelékek fejlődését, amelyek aztán nagyon különböző struktúrákká differenciálódnak a különböző törzsekben.
Az egyik leghíresebb példa az ilyen génekre az eyeless, amelyet eredetileg egy kisebb szemekkel bíró, vagy egyenesen szemnélküli, mára klasszikus Drosophila mutánsban azonosítottak. Az eyeless emlős homológjának, a Pax6-nak a mutációja szintén a szem leépüléséhez vezet. A Pax6 és eyeless a szem fejlődését és differenciálódását szabályozó fehérjék egy hierarchiájának átírását aktiválja. Mindkét gén kifejeződése a fejlődő szemkezdeményre korlátozódik. Amikor egy csodálatos kísérletben a kutatók elérték, hogy az eyeless gén a testnek olyan részeiben is kifejeződjön, ahol egyébként néma marad, azaz ektopikusan is megjelenjen a géntermék, az eredmény minden elképzelést felülmúlt: kiderült, hogy ez az egyetlen gén elégséges ahhoz, hogy indukálja ektopikus szemek megjelenését ott, ahol kifejeződik. Talán még sokkolóbb a Pax/eyeless gén konzerváltsága: a tintahal és az egér Pax6 génje képes a szem idukciójára Drosophilában! A további kutatások szerint a Pax6 által szabályozott géneknek legalábbis jelentős része szintén konzerválódott, ami magyarázza, hogy miért képes a gén azonnal irányítani a fejlődést egy ennyire különböző genomban.
A szem komplexitása mióta világ a világ foglalkoztatja a kutatókat, komplexitása, „összecsiszoltsága” mindig is téma volt. Darwin arra a következtetésre jutott – megkönnyebbülve, hogy nem kell „sutba vágnia” elméletét a szem miatt sem -, hogy különböző átmeneti állapotú érzékszerveknek, amelyek képesek a fény érzékelésére, van adaptív értékük, és rajtuk keresztül vezethetett az evolúció egy olyan komplex szervhez, mint a szem. Ezzel az elgondolással egybevág, hogy a többi törzstől legrégebben elvált, ún. bazális törzseknél, amelyek – jó közelítésben - ősi állapotot tükröznek, néhány sejtből álló, fényérzékeny pigmentet (rodopszint) tartalmazó „szemfoltokkal” rendelkeznek, és számos szemtípus van jelen az állatvilágban. Amellett, hogy a szemek több tucatszor kialakulhattak, a Pax6 és célgénjei közös eredetre utalnak. Az elfogadott érvelés szerint a Pax6 nagyon ősi funkciója, hogy szabályozza a fotoreceptor szervek univerzális komponenseinek kifejeződését, de a különböző állati vonalakban a különböző morfológiai sajátságok aztán függetlenül fejlődtek ki olyan gének irányításával, amelyek „alárendelődtek” a Pax6 útvonalnak. A képesség a fotorecepcióra ezek szerint homológ sajátság, és a konzerválódott ősi elemekre szokás a „mély homológia” („deep homology”) kifejezést használni, de a kivitelezésben részt vevő strukturális elemek, azaz a szemek, a szó klasszikus értelmében legalábbis, nem homológok, számukra talán megtarthatjuk az analóg kifejezést. A többi útvonalakra is minden bizonnyal hasonló elvek érvényesülnek.
A Drosophila homeotikus génjeinek klónozása után hamarosan következett a XX. század egyik leginkább forradalmi tudományos felfedezése: MINDEN ÁLLATI GENOMBAN VANNAK EZEKKEL ROKON GÉNEK
(módszer: a homeobox, ez a 180 bp hosszúságú DNS-szakasz, amely minden Drosophila HOX-génben megvolt, próbaként felhasználva hibridizált más élőlények génjeihez, amelyek gyakorlatilag ugyanezt a szakaszt tartalmazták)
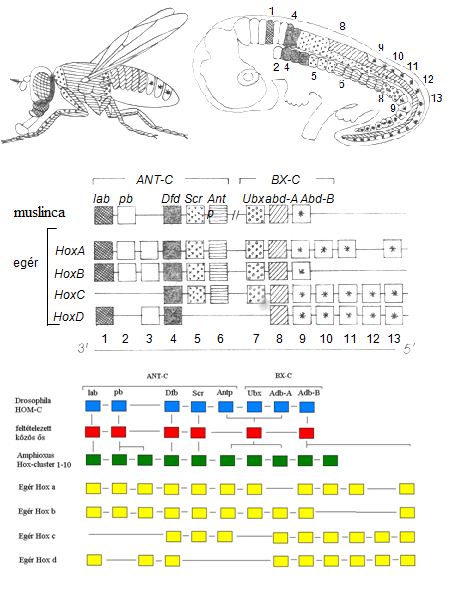
Meglepő eredmény: A gerincesek Hox-génjeinek fizikai térképezése felfedte, hogy 4 nagy komplexben helyezkednek el, a komplexeken belül a gének sorrendje megfelel a leginkább rokon (leginkább homológ) Drosophila génekének
A Hox-gének kifejeződési rendje az anteroposzteriorális/rosztrokaudális tengely mentén megfelel a gének pozíciójának a génklaszterben
(a klaszterekben nem ugyanannyi gén van, de logikájuk ugyanaz) 20. ábra
10.20. ábra - A homeotikus gének szerveződésének, tér-és időbeli kifejeződési mintázatának konzerváltsága Drosophilában és egérben. A HOM-C, az Antennapedia- és bithorax-komplexek összességét jelöli a Drosophila harmadik kromoszómáján. Az egér a HOM-C génjeinek megfelelő Hox génjei négy klaszterben – HoxA, HoxB, HoxC, HoxD – helyezkednek el. Az egymásnak homológia alapján megfelelő géneket azonos színezéssel jelöltük. Minden egyes egér Hox gén azonos irányban íródik át (az egyes géneknél az átíródás iránya 5'®3'). Az egyed szintjén minél előbbre fejeződik ki a gén, annál korábbi is annak kifejeződése. Egérnél bemutatjuk a gének kifejeződését a központi idegrendszerben és a szomitákban. Bizonyos kifejeződési domének átfednek. Gyakorta csakis az elülső kifejeződési határ éles
 |