Mezők, kompartmentek, organizátorok- a szelektor gének expressziós doménjai vagy jelmolekulák forrásai- molekuláris bizonyíték alapvető szervező szerepük mellett
Toolkit konzerváltsága: számos kérdést vet fel
-
-
Hogyan irányíthatják ilyen hasonló gének ilyen különböző szervek fejlődését, mint az összetett-, és a hólyagszem?
-
Mit mond a hasonló funkciójú, de oly különböző struktúrák kialakításában fontos gének struktúráltsága közös ősünkről?
-
A „fejlődési” gének azonosítása csak az első, bár igen fontos lépés a fejlődésbiológiai analízisben. A fő kérdés: HOGYAN BONTAKOZIK KI AKTIVITÁSUK TÉRBEN ÉS IDŐBEN, MIKÖZBEN IRÁNYÍTJÁK A TESTTERV ÉS A TESTRÉSZEK KIALAKULÁSÁT?
A fejlődésben a GÉNSZABÁLYOZÁS A SAROKPONT - az EPIGENETIKA elmélete foglalja keretbe a fejlődéssel kapcsolatos génszabályozást.
MI IS AZ EPIGENETIKA?
Új elmélet, amely a sejtet tekinti alapvető egységnek, és az abban lejátszódó génkifejeződéssel kapcsolatos folyamatokat a fejlődés hajtóerejének. A génkifejeződés változásait tanulmányozza a differenciált sejteket tartalmazó élőlényekben, valamint adott génkifejeződési mintázatok továbböröklődését a mitotikus osztódások folyamán. Foglalkozik továbbá olyan, sejtmagi öröklődéssel is, amely nem a DNS-szekvencia megváltozásán alapul. Az epigenetikai szabályozás azoknak a genetikai faktoroknak az összessége, amelyek a sejtre hatva szelektíven szabályozzák a génkifejeződést, ami az adott sejt epigenotípusát adja, és a fejlődés folyamán az egyre összetettebb fenotípusokhoz vezetnek
Az epigenetika és a differenciális génexpresszió tehát erőteljesen átfedő fogalmak. Két különleges esetet a teljesség igény nélkül megemlítünk. Az epigenetikai szabályozás sokszor a DNS metilációs mintázatának megváltozásán alapul: A genomi imprintingnél az apai és anyai genom bizonyos, eltérő részei (metiláción alapuló) kromatinszerkezetet vesznek fel és inaktiválódnak és ez a mintázat az életen át megmarad. Ha az inaktiváció mindkét (apai és anyai) genomban megtörténik vagy mindkettőben teljesen elmarad, súlyos fejlődési következmények lépnek fel. Tehát a szekvencia lehet ugyanaz az apai és anyai genomban, a gének kifejeződése mégis szélsőségesen különbözhet az imprintinget mutató régióban.
M ásik példa az X-inaktiváció emlősökben. A nőstény emlősökben a fejlődés korai stádiumában minden sejt egyik X kromoszómája véletlenszerűen inaktiválódik és ez az állapot fennáll az egész élet folyamán.
Promóterek és enhenszerek
Kétféle típusú szabályozó elem szükséges ahhoz, hogy a gének a megfelelő helyen és időben íródjanak át. Az egyik típust cisz-regulátor elemnek nevezzük. Specifikus DNS-szekvenciákat jelent meghatározott kromoszómákon. Csakis a szomszédos génekre fejti ki hatását (leszámítva néhány, igazán speciális esetet).
Eukarióta fehérjét kódoló (pol II) géneknél két cisz-regulátor elem befolyásolja a transzkripció idő- és térbeli specifitását: az enhanszerek és a promóterek.
A promóterek közvetlenül 5’ irányban, (= upstream = folyásirányban felfelé jelentésű kifejezés; sokszor használják olyan szabályozóelemekre, amelyek 5’ irányban helyezkednek el az átírt RNS szempontjából, vagy bármilyen hierarchiában feljebb állnak a kérdéses elemnél), helyezkednek el azoktól a helyektől, ahol a transzkripció iniciálódik. Általában több száz bázispár (bp) hosszúságúak.
A cisz-regulátorok másik típusa az enhenszerek. A hetvenes évek végén fedezték fel, hogy az eukariótákban vannak olyan DNS szekvenciák, amelyek sok ezer bázispárnyi távolságról is képesek aktiválni a transzkripciót bizonyos promóterekről. Ma már ismert, hogy ezek az ún. enhenszer szekvenciák specifikus kötőhelyként szolgálnak a génszabályozó fehérjék számára, melyek elősegítik, növelik (angolul: enhance) a transzkripciót. Ez a típusú távolba hatás általánosnak bizonyult az eukariótáknál. A távolhatás megmagyarázható a kihurkolási modell segítségével, melynek a lényege az, hogy a promóter és enhenszer közötti DNS-szakasz kihurkolódik, és a hozzájuk kapcsolódó fehérjék közvetlenül érintkezhetnek (a modell megalkotásában magyar kutatónak, Orosz Lászlónak is nagy része van; 6. ábra). Az enhancer-DNS e szerint a felfogás szerint „gyűjtőhelyként” szolgál az enhenszerhez kötődő fehérjék számára, és elősegíti ütközésüket a promóter fehérjéivel. (Hasonló hatás érvényesülne az adott fehérjék promóter körüli helyi koncentrációjának emelésével.)
10.6. ábra - A génkifejeződést a DNS szabályozó régióihoz kötődő génszabályozó fehérjék koordinált tevékenysége szabályozza. A transzkripciós apparátus – amelyet az RNS polimeráz II és az általános transzkripciós faktorok alkotnak – a promóterhez kötődik. A további szabályozó régiók, amelyekhez génszabályozó fehérjék kötődnek, elhelyezkedhetnek a promóter mellett, vagy nagyobb távolságban
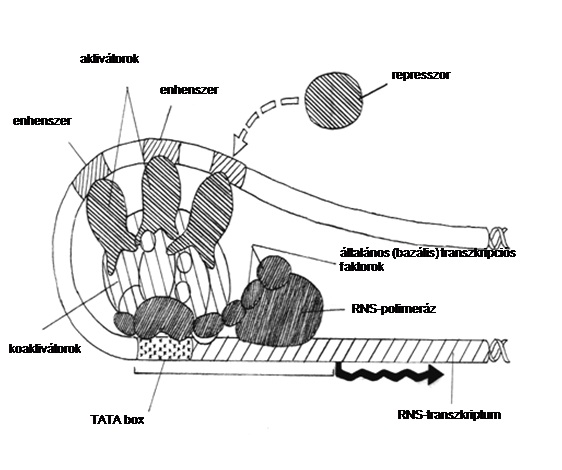 |
Az enhenszerek (enhancer) elősegítik a promóterek működését. Szabályozzák a róluk történő transzkripció hatékonyságát és mértékét. Az enhenszerek cisz-helyzetű promóterekkel vannak kölcsönhatásban, de távolságuk azoktól nagyon nagy, akár 50 kb is lehet (írtak le olyan esetet, amikor nem cisz helyzetű, azaz más-más kromoszómán lévő promóter és enhenszer kölcsönhatása bizonyosnak látszik, és jóval nagyobb távolságokra való hatást is, de most az általános esetre összpontosítunk). Elhelyezkedhetnek 5’ vagy 3’ irányban a promóterhez viszonyítva, vagy akár intronokban is lehetnek. Az enhenszerek – a promóterekhez hasonlóan – transzkripciós faktorokat, transzregulátorokat kötnek meg, azokon keresztül fejtik ki hatásukat. Újabban szokásos a promótert és enhenszert együttesen „promóternek” tekinteni, ilyenkor az előbbi, klasszikus értelemben vett promótert „mag (core) promóter”-nek nevezik.
Az enhenszerek egy sajátos típusa a negatív enhenszer, vagy szájlenszer (silencer). Ha a megfelelő transzkripciós faktorok kötnek a szájlenszerekhez, azok megakadályozzák a cisz-elhelyezkedő promóterekről történő átírást. A szájlenszerekhez kötődő fehérjék kötődése speciális kromatinstruktúrához vezet. Itt megjegyezzük, hogy a génekrepresszió „szájlenszing”-nek (silencing) nevezett módja nem mindig klasszikus szájlenszerekkel kapcsolatos.
Az enhenszerek elsődleges funkciója annak a repressziós, gátló hatásnak a megakadályozása, amely a kromatinstruktúra bizonyos komponenseitől ered, más oldalról nézve a kromatin aktiválása.
A transzkripciós faktorok enhenszerekhez kötődése az esetek legalábbis nagy részében előfeltétele az általános transzkripciós apparátus működésének.
Az enhenszerek meghatározóak a fejlődés szabályozásában. Vizsgálatuk néhány általános vonásukra hívja fel a figyelmet, amelyek a differenciális génexpresszió szempontjából fontosak.
-
A legtöbb gén szabályozásában részt vesznek.
-
Az enhenszerek a fő meghatározói a differenciális transzkipció tér- és időbeliségének.
-
Egy génnek számos enhenszere lehet, és minden egyes enhenszerhez kötődhet több faktor is. Így esetleg működhetnek enhenszerként és szájlenszerként – vagyis bizonyos gének kifejeződését negatívan befolyásoló szabályozóelemekként – is.
Transz regulátorok
A szabályozó faktorok másik csoportját transz-regulátoroknak nevezzük. Szolubilis, azaz nem helyhez kötött molekulák, amelyek kölcsönhatásba lépnek más kromoszómális lokalizációjú génekkel. Lehetnek fehérjék vagy bizonyos speciális, de nagyon fontos esetekben RNS-ek. Fejezetünkben őket, erősen vitathatóan, elhanyagoljuk, a sejtmagban történtekre (és a sejtmagba vezető útra) összpontosítunk
A transzkripciós faktorok olyan fehérjék (transz regulátorok), amelyek az enhenszer/szájlenszer és a promóter régiókhoz kötődnek. 7. ábra
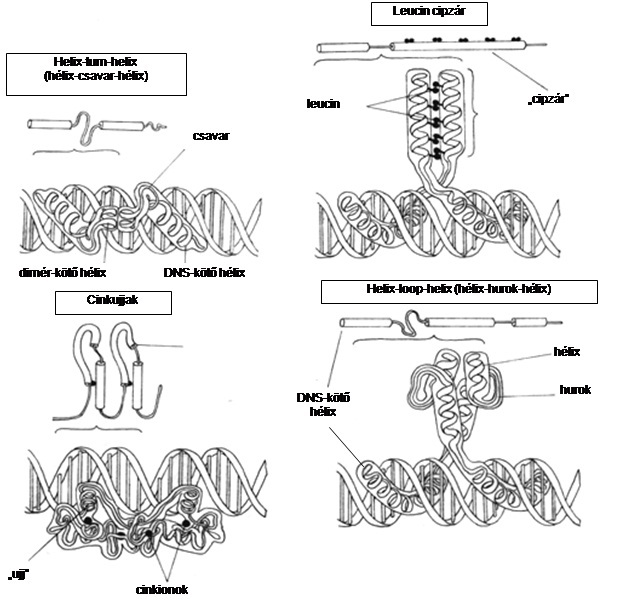
A legtöbb transzkripciós faktor specifikus, a szabályozó régiókon belül is jól körülírt DNS-szekvenciákhoz kötődik. Szokás őket kötődésük alapján családokba sorolni. Családon belül a fehérjék DNS-kötő régiójának a szerkezete hasonló, de a régió aminosavai közötti kis különbségek, ill. más faktorokkal való kölcsönhatásuk oda vezethet, hogy a nagyon közeli rokon fehérjék is más DNS-szekvenciákhoz kötődhetnek.
Fontos tudatosítanunk, hogy a DNS egy szakaszának háromdimenziós szerkezete a transzkripciós faktorok számára használható információkat rejt, és ezt a tényt a génexpressziót szabályozó fehérjék célszekvenciáik keresésekor ki is használják.
Összeáll a rendszer - megindul a fehérjeszintézis
Az mRNS szintézise bonyolult fehérjekomplexumok létrejöttét kívánja meg az eukarióta gének promóterén.
A folyamathoz két, már említett különböző DNS-elem jelenlétére van szükség: a szűk értelemben vett (core) promóterre és enhenszerekre.
A promóter meghatározza a transzkripció iniciációjának pontos helyét, míg az enhenszerek irányítják a transzkripció térbeni és időbeni mintázatát. A promóter egyben a bazális transzkripciós faktorok alkotta általános transzkripciós „gépezet” horgonyzási helye is.
A promóterrel szemben az enhenszer elemekhez gén-specifikus aktivátorok kötnek, amelyek serkentik az átíródást. A transzkripciós aktivátorok hiányában in vitro az eukarióta gének gyengén, „alapszinten” (basal level) íródnak át.
A cisz-regulátor elemek felépítése
A promóterek és enhenszerek sok esetben lehetnek moduláris természetűek, ami azt jelenti, hogy az általuk szabályozott gén bekapcsolásához egy adott helyen nem az egész promóter- vagy enhenszerrégióra, hanem csak egyes modulokra van szükség. Az egyes modulok aktivitása attól függ, hogy bizonyos, hozzájuk kötődő transzkripciós faktorok jelen vannak-e az adott sejtben. Ha igen, a kérdéses sejtben a gén aktív lesz. Más sejtekben más modulok aktiválhatják a gént, így áll össze a teljes kifejeződési mintázat. A modulos szerkezet nagy rugalmasságot biztosít a gén kifejeződésében. A transzkripciós faktorok aktivitásának szabályozásánál, ill. több fejlődési folyamatnál, konkrét példát is láthatunk a moduláris szabályozásra
-
A METAZOÁK CISZ-REGULÁTOR DNS-ÉNEK MODULÁRIS FELÉPÍTÉSE ELSŐDLEGES FONTOSSÁGÚ A FEJLŐDÉS ALATTI GÉNKÖLCSÖNHATÁSOKBAN ÉS AZ ÚJ MORFOLÓGIÁJÚ ÉLŐLÉNYEK KIALAKULÁSÁBAN.
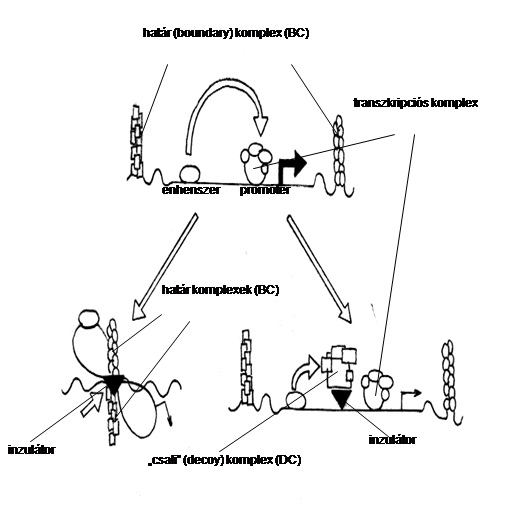
A magas szintű átiródáshoz az enhenszerek és promóterek közötti kölcsönhatásra van szükség. A „doménhatár” modell szerint az inzulátoron kialakuló fehérjekomplex (magasabb szerveződésű kromatinstruktúra) fizikailag választja el a promótert és az enhenszert, lehetetlenné téve kommunikációjukat. 8. ábra.
10.8. ábra - Boundary vagy inzulátor régió működésének modelljei:A „transzkripciós csalétek” (transcription decoy) modell szerint az inzulátoron olyan fehérjekomplex szerveződik, amely hasonlít a transzkripciós apparátusra. Az enhenszer így a „hamis” komplexszel lép kölcsönhatásba, és nem segíti elő a hatékonyabb transzkripciót
 |
Egy struktúrára, mintázatra, vagy folyamatra sok gén gyakorolhat kisebb-nagyobb hatást. A mutáns fenotípus tanulmányozása sokszor nem ad támpontot a gének kifejeződési sorrendjére vonatkozóan, és általában az sem világos pusztán a fenotípus alapján, hogy melyik gén játszik elsődleges, melyik alárendelt szerepet.
A végleges forma helyett a génkifejeződési mintázatokat használják - közvetlen és dinamikus képet adnak a génfunkcióról és a regulatív kapcsolatokról.
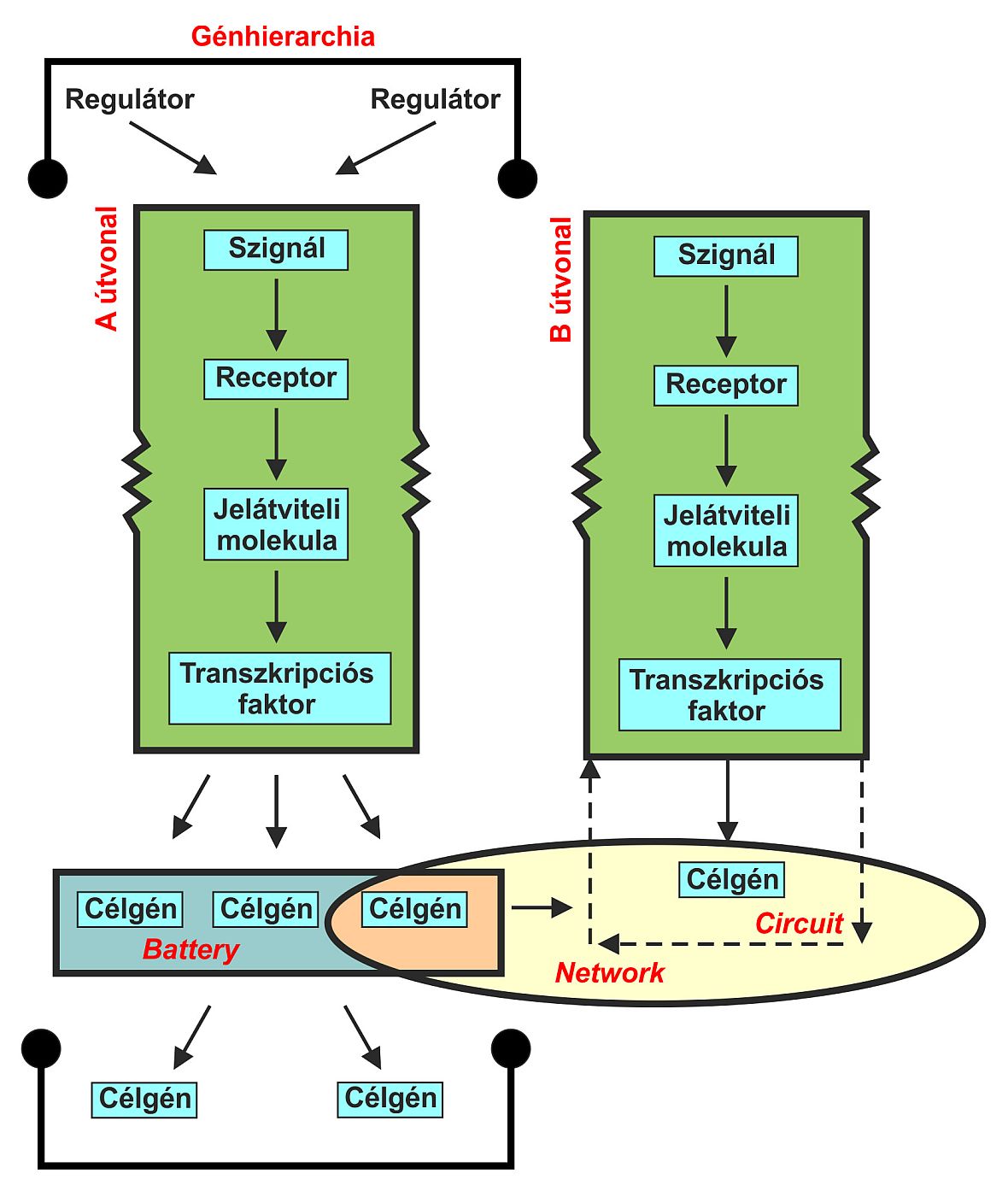
A génszabályozási hierarchiák általános sajátságai:
1. a fejlődés egy kontinuum, amelyben minden génkifejeződési mintázatnak megvan az előzménye: egy másik génkifejeződési mintázat
2. a fejlődést szabályozó információk gyakorta folynak át „csomópontokon”, vagyis kulcsgéneken, amelyeknek kifejeződése döntően befolyásolja a későbbi mintázatot
3. sok gén a fejlődés több stádiumában aktív (jelátviteli gének, transzkripciós faktorok) – több hierarchiában is részt vesznek
Pathway - szükségszerűen összekötött elemek az információáramlásban, pl. jelátviteli útvonalak
Circuit - nagyobb, mint a pathway, elemei nem kötelezően kapcsoltak. Olyan szabályozó útvonal, amelyhez bizonyos target gének tartoznak.
Összekötött circuit-ok network-öt alkotnak. Némely gének korábban hathatnak, vagy más géneket szabályozhatnak, ezek hierarchiává szervezik a network-öt. A hierarchia szerveződhet vertikálisan („tier”-ek), vagy horizontálisabban, ilyenkor egyes gének közvetlenül szabályozzák a target gének nagy csoportját (battery) 9. ábra