A „koevolúció” fogalom természetesen alkalmazható a fajok párhuzamosan zajló diverzifikációjára, amelyet a társas viszonyban lévő fajok filogenetikai viszonyai is tükröznek. Szépen párhuzamba állítható („konkordáns”) filogenetikai fáik vannak a gazdafajoknak parazitáikkal és endoszimbiontáikkal (pl. a méheknek az őket triptofánnal ellátó Buchnera baktériumokkal). A feltevések szerint a méhek már a kezdeteknél fertőzöttek voltak a baktériumokkal, majd a fajok elválása után a baktériumok együtt divergálódtak „új méhfajukkal”, a horizontális transzfer, azaz fajok közti baktériumcsere, vagy gazdacsere (host switching) pedig gyakorlatilag nem történt, ami talán az endoszimbionta baktérium és a méhek a szokottnál is intimebb viszonyának köszönhető.
Ha a parazita a környezeten át terjed egyik gazdáról a másikra, mint a növényevő rovarok, nagyobb valószínűséggel váltanak gazdanövényt, így a filogenetikai mintázatok a növények és növényevők között ritkán mutatnak erős konkordanciát. A filogenetikai viszonyok így is utalhatnak ősi kapcsolatokra: a levélbogarak, cincérek és az ormányos bogarak tápláléknövényei főleg cikászok és fenyők, míg a „fejlettebb” bogarak zárvatermőkkel táplálkoznak. Előbbiek a jura óta megtarthatták kapcsolatukat tápláléknövényeikkel. A régi szép kapcsolatoknak néha kézzelfogható nyomai is vannak: a mai gyömbérnövényen ugyanolyan rágásnyomok vannak, mint krétakori elődjén.
A ragadozóknak és a parazitáknak számos, igazán látványos adaptációja alakult ki áldozatuk legyűrésére. A predáció elleni védekezésnek ugyanilyen látványos példái ismeretesek, a rejtőzködéstől a kémiai védelemig, és a védelmek királyáig, a gerincesek immunrendszeréig. Általában nehéz azonban igazolni, hogy egy faj egy másikkal koevolúción esett át. Elméletben a ragadozó és préda viszonya többféleképpen alakulhat:
- Folytatódhat a végtelenségig az evolúciós fegyverkezési harc eszkalációjával
- Létrejöhet szilárd genetikai egyensúly
- Kialakulhatnak folyamatos ciklusok, vagy szabálytalan fluktuációk a két faj genetikai összetételében
- Vezethet egyik vagy mindkét faj kihalásához
A végtelenbe nyúló fegyverkezési verseny nem valószínű, mert a támadó- vagy védekező kapacitás fejlesztése idővel olyan energiaráfordítást kíván, hogy a költség meghaladja a hasznot. A tölgyek tanninjának vagy a fenyők terpénjeinek termelése a növény energiatermelésének 10 százalékát felemészti, a pasztinák (Pastinaca sativa) a kertben magasabb furanokumarin tartalma miatt védettebb a szövőlepke-hernyók ellen, és több magvat termel, míg ugyanezek a növények az üvegházban, védett környezetben kevesebb magvat termelnek kisebb furokumarin tartalmú társaiknál. Így érthető, hogy a növények teljes védettséget nem érnek el a növényevők ellen.
Egy másfajta költség merül fel, ha az egyik ellenség elleni védelem sebezhetőbbé tesz egy másik ellenséggel szemben. Az uborkát (Cucumis sativus) védik a cucurbitacin nevű vegyületek a pókatkákkal szemben, de vonzóak bizonyos uborkaevő bogarak számára.
A természetes populációkra hosszú távú koevolúciójára vonatkozóan, de közvetett bizonyíték nagy számban áll rendelkezésre. A mezozoikumban új, nagyon effektív puhatestűekre specializálódott ragadozók fejlődtek ki, elsősorban a héjat szétzúzni képes halak és rákfélék. A csigák és kagylók diverzitása is megnőtt, vastagabb héjak, kisebb és védettebb bejáratok, tüskék, és más képletek alakultak ki a ragadozók elleni védelem érdekében, és a regenerálódási képesség is fejlődött bizonyos vonalakban.
Az Amerika észak-nyugati részén élő Taricha granulosa nevű érdes bőrű gőte rendelkezik az egyik leghatékonyabb védelmi rendszerrel a ragadozók ellen: legtöbb populációjának nagy mennyiségű tetrodotoxin (TTX) van a bőrében, ami félelmetesen hatásos idegméreg (egy példánynak 25 000 egér megöléséhez elégséges mennyiség). A Thamnophis sirtalis kígyónak azok a populációi, amelyek azonos területen élnek (szümpatrikusak) a mérget termelő gőtékkel, százszor rezisztensebbek a méreganyagra, mint az allopatrikus populációk. 2. ábra

A paraziták két nagy kihívással szembesülnek: hogyan juttatják át magukat és utódaikat egyik gazdáról a másikra (transzmisszió), és hogyan győzik le a gazda védelmi rendszerét. Néhány parazita, mint a rovarpetékben továbbadódó Wolbachia vertikálisan adódik át, a szülőről utódjára, mások horizontálisan adódnak át a populáció tagjai között, a külső környezet közvetítésével, kontaktussal, vagy vektorral.
Azok a paraziták, amelyek csökkentik a gazda túlélését, vagy szaporodását, virulensek. Elsődleges cél azoknak az evolúciós faktoroknak a megismerése, amelyek a virulencia mértékét befolyásolják. A virulencia mértékének változása lehet nagyon gyors a „mikroparazitáknál”, a vírusoknál és baktériumoknál, így a kérdésnek nagyon nagy az orvosi jelentősége is. A virulencia szintje függ a gazda és a parazita evolúciójától is. Klasszikus példa, hogy amikor az üregi nyulat (Oryctolagus cuniculus) betelepítették Ausztráliába, komolya kártevővé vált. Védekezésül dél-amerikai nyulak myxoma vírusait telepítették be. Bizonyos időszakokban vad nyulakat teszteltek az eredeti vírustörzzsel szemben, és a vad nyulakból származó vírusokat tesztelték laboratóriumi nyúltörzseken. Az idők folyamán a vadon élő nyulak nagyobb rezisztenciát fejlesztettek ki a vírussal szemben, míg a vírusnál alacsonyabb virulancia alakult ki. Teljesen avirulens vírustörzs nem alakult ki.
Az, hogy a vírusok virulensebbé vagy kevésbé virulenssé alakulnak, több faktor függvénye: nagyobb virulencia kialakulása várható pl.
-
olyan parazitáknál, amelyeknél a többszörös fertőzés esélye nagyobb (a nagyobb reprodukciós rátájú vírus szelektálódhat),
-
az olyan gazdánál, amely hamarabb válik immunissá (nagyobb szaporodási rátájú vírusok szelektálódnak,
-
amelyek „túlszaladnak” az immunrendszer védelmi kapacitásán),
-
és a horizontális terjedés esetén (ilyenkor a gazda szaporodása nem feltétele a továbbadódósnak, a vírusnak nem „érdeke” annak kímélése).
A Daphnia magna parazitája egy mikrospórákat termelő (Pleistophora intestinalis), amely a bél epitéliumban szaporodik, és spóráit az ürülékbe ereszti. Fertőzött, és fertőzetlen Daphniákat hoztak össze. Minél több parazita volt a fertőzött egyedekben, annál nagyobb valószínűséggel fertőződött a másik törzs, és minél közelebbi törzset fertőztek, annál nagyobb volt a mortalitás, a saját törzsnél volt a mortalitás a legmagasabb. A paraziták tehát semmiképpen nem törvényszerűen „szelídülnek” evolúciójuk során.
Dybdahl és Lively (1998) a Potamopyrgus antipodarum édesvízi csiga és közvetett fejlődésű métely (Trematoda) parazitáját (Microphallus sp.) vizsgálta. A csigapopulációban szexuális és szűznemzéssel szaporodó egyedek egyaránt jelen voltak. Ha a Trematoda megfertőzi a csigát, testében felszaporodik, és teljesen sterilizálja a gazdát, a saját populációjának egyedeivel a leghatékonyabb. A leginkább aszexuális csigákkal belakott tó különböző klonális fenotípusainak gyakorisága (izoenzim markerek alapján), 5 év alatt megváltozott: a legtöbb genotípus gyakorisági maximuma utáni évben infekciós maximuma is elkövetkezett. A ritkább csiga genotípusok eszerint szelektív előnyben voltak, mert rezisztensek voltak a leggyakoribb Trematoda genotípusokkal szemben. Ahogy a csiga genotípusok gyakorisága változott, aztán elveszítették előnyüket. A hipotézist úgy tesztelték, hogy 40 ritka csigaklónt és 4, a legutóbbi időben gyakori csigaklónt fertőztek, és az előbbiek csakugyan kevésbé voltak érzékenyek a fertőzésre. A szexualitás szerepe itt tetten érhető: új, ritka, rezisztens genotípusok csak szexuális úton jönnek létre.
A mutualizmus olyan, fajok közötti kölcsönhatás, amelyből mindegyik félnek haszna van. A szimbiotikus mutualizmuskor a különböző fajokba tartozó egyedek életük nagy része alatt meghitt közelségben vannak. Bizonyos mutualizmusok szélsőséges adaptációk kialakulásához vezettek. Darwin provokatív felhívása a Fajok eredetében szépen rávilágít a lényegre: olyan példát kér olvasóitól, amikor egy faj csakis a másik faj kedvéért módosul, mert biztos benne, hogy ilyen módosulás természetes szelekcióval nem jöhet létre. A mutualizmus nem altruizmus, hanem a az előnyök kölcsönös kiaknázása, akkor is, ha bizonyos mutualizmusok visszavezethetők parazitikus, vagy egyéb, a másik fajt kizsákmányoló kapcsolatokra. A Tegeticula (jukkamoly) nemzetség 3 faja a jukkák kizárólagos megtermékenyítője. A molyok csak jukkával táplálkoznak. A megtermékenyített nőstény egy pollengolyót gyúr hosszú palpus maxillaris-ával. A golyóval egy másik növényre repül és egy fiatal virágot választ.
A termő magházának mindhárom kompartmentjére petét rak. Epideiktikus feromonok, melyek az egyedek közötti „távolságtartást” segítik elő, megakadályozzák, hogy a magházra túl sok pete kerüljön. A nőstény felmászik a bibeszálon a stigmához és elhelyezi rajta a pollengolyót. A lárvák a fejlődő magkezdeményt fogyasztják. A talajban bábozódik. 3. ábra
8.3. ábra - Tegeticula yuccasella fajkomplex nem minden esetben pollinátor és néha magára a termésre petézik
 |
A koevolúció eredménye:
A Tegeticula oldaláról: - pollengolyó gyúrása - a golyó elhelyezése a stigmán - obligát oligofágia - 3-45%-nál több ováriumra nem petézik - a virággal szinkronizált kelés miatt a bábok 1-3 évig fekszenek
A Jukka oldaláról: - kizárólag Tegeticulák porozzák be - a 6-nál több lárvát tartalmazó magházak abortálódnak - a petézésre csak 5 nap áll rendelkezésre - a virágzás néha elmarad - a beporzott termőknek csak 6 %-ából fejlődik termés
Tegeticula yuccasella fajkomplex nem minden esetben pollinátor és néha magára a termésre petézik Az evolúciós stabilitás kialakulására is szép példát adnak a jukka fajok és az őket kizárólagosan megporzó molylepkék tehát minden egyes virágba csak néhány petét raknak, hogy a sok fejlődő mag közül csak néhányat fogyasszon el a fejlődő lárva. Miután petéit több virág között osztja el, miért nem rak többet egy virágba? Biztosan fontos, hogy a növénynek egyszerűen nincsen elég forrása ahhoz, hogy 500-1500 virágjából gyümölcsöt érleljen. Amikor a kutatók kézzel megporozták néhány növénynek minden virágját, azt tapasztalták, hogy mindössze a virágok 15 százaléka érett magvas gyümölccsé, a többi abortálódott, és leesett a növényről. A további vizsgálatok megmutatták, hogy a több petét tartalmazó virágok nagyobb eséllyel abortálódnak, azaz nagy a szelekciós nyomás a túl sok petét rakó egyedeken.
6.4.6 A fajok: Ficus sycomorus és a Ceratosolen arabicus (Agaonidae) - a növény egylaki, a virágok egyivarúak. - A női virág lehet hosszú és rövid bibeszálú. - lamellás zárószerkezet - kivezető nyílás (ez hiányozhat is) 1. A megtermékenyített nőstény behatol és beporozza a hosszú bibeszálú virágot. 2. Petét rak a rövid bibés virágokra, majd elpusztul. 3. A virágokból gubacs fejlődik, melyekből a kikelő lárvák táplálkoznak. 4. A hím darazsak korábban kikelnek. 5. A hímek a gubacsban megtermékenyítik a még bent levő nőstényeket, majd elpusztulnak (ha van kivezető nyílás). Ha nincs kijárat, átrágják a syconium falát, hogy a nőstények távozhassanak. 6. A nőstények virágport gyűjtenek és a kijáraton át távoznak. Minden fügefajnak „saját” fügedarázsfaja van, mely csak az adott növényen képes szaporodni. Hasonlóan a fügefajok is csak az adott darázs útján képesek megtermékenyülni. Sok olyan adaptáció figyelhető meg (főként a darazsakon), melyek a koevolúció következményei, pl. a könnyebb közlekedés céljából a nőstények a fügébe hatolás közben ledobják szárnyaikat; a nőstény a rövid bibés virágokra petézik, a hímek a gubacsban termékenyítik meg a szűz nőstényeket. További igen érdekes részletekkel szolgál Dawkins: A Valószínűtlenség Hegyének meghódítása c. könyve. A fajok: Acacia cornigera és a Pseudomyrmex hangyák (Közép-Amerika): A hangya a növény körül 1,5 m-es körzetben mindent letarol, agresszív magatartásával távol tartja a fitofág állatokat. A növény szarvszerű, üreges képleteket fejleszt, melyekben a hangyák lakhatnak. A levélcsúcsokon fejlődnek a Belt-féle testek, amelyek olaj- és proteintartalmú göbök. Ezekkel táplálkoznak a hangyák.


4. ábra Ha mutualizmus alakul ki, mindig potenciális veszélyt jelent, hogy felbukkan egy genotípus, amelyik „csal” (angolul: „cheats”), azaz kiaknázza a partner által nyújtott előnyöket, anélkül, hogy megfizetné az árat, vagyis biztosítaná az előnyt partnere számára. A „csalónak” szelektív előnye van, amíg nem túl gyakori, és nem alakul ki ellene védelem. A szelekció előnyben részesíti azokat a legalább egyik fajban fellépő mechanizmusokat, amelyek megakadályozzák a túlzott kiaknázást, sőt, a szelekció előnyben részesíti a „becsületes” genotípust, ha az egyed genetikai érdeke a gazda vagy a partner jó erőnléte. A mutualizmus evolúciós stabilitását elősegítő tényezők lehetnek: az endoszimbionták szülőkről utódokra való továbbadódása (vertikális transzfer), ismételt vagy élethosszig tartó társulás ugyanazzal a partnerral vagy gazdával, korlátozott lehetőség más partnerre való váltásra, vagy más források együttes felhasználása. A méhek és Buchnerák mutualizmusa kielégíti a fenti elveket: Minden, a méhek sejtjeiben élő Buchnera vertikálisan öröklődik, és valamennyi ismert faj mutualisztikus. ( Vannak emellett vertikálisan öröklődő, veszélyes szimbionták is.) A mutualizmus evolúciós időskálán nem mindig stabil: sok faj csal. Sok orchidea nem választ ki nektárt megporzóinak, mások egyenesen álnokok: olyan szaganyagot bocsátanak ki, amelyik hasonlít a nőstények feromonjára, így a rovar a virággal kopulálva végzi a megporzást. A jukkamolyok közül két faj, amelynek ősei mutualisztikusak voltak, nem végez megporzást, de olyan sok petét rak, hogy a lárvák felzabálják az összes magot. A növény abortáló válaszát a túl sok petére úgy játsszak ki, hogy az után a periódus után rakják le petéiket, hogy a virágok elabortálódása bekövetkezhet.
Ha két faj ugyanazokat a forrásokat használja, közöttük versengés alakul ki (interspecifikus kompetíció). Darwin posztulálta, hogy a versengés következménye a források felhasználása terén diverzifikálódáshoz vezető szelekció, amit a fajok eredetének és diverzifikációjának fő okának tekintett. Ma már bizonyítottnak tekinthető, hogy a versengés hatására bekövetkező evolúciós változások az adaptív radiáció legfőbb okai. Az ökológiai kutatások kimutatták az azonos élőhelyen élő (szimpatrikus) fajok jellemző módon különböznek a források felhasználásában. Legalábbis plauzibilis feltételezni, hogy ezek a különbségek, legalábbis részben, a kompetíció elkerülésére alakultak ki. Magevő madaraknál pl. könnyű belátni, hogy a szélsőségesebb csőrfenotípussal rendelkező egyedek kevésbé kényszerülnek versenyre fajuk és más fajok egyedeivel, ha a magvak egy széles nagyságspektruma áll rendelkezésre. Így a legextrémebb fenotípusok (genotípusok) lesznek a legsikeresebbek: ez a denzitásfüggő diverzifikáló szelekció eredményezhet két új fajt, amelyek kevésbé fednek át a források felhasználásában, és fenotípuseloszlásuk is különböző lesz. A fajok közötti versengés hatására létrejövő divergenciát gyakorta nevezik ökológiai jellegeltolódásnak (ecological character displacement). Mivel a lókuszok közötti rekombináció korlátozza a források felhasználásában meghatározó poligénes jellegek varianciáját, sokszor egy-két faj nem képes lefedni a források széles spektrumát. Ilyen esetekben egy vagy több, a már jelenlévőktől független faj adódhat a közösséghez, és az összes, jelenlévő faj evolúciója tovább finomítja a források felhasználását, hogy a versengés minimalizálódjon. A fajok aztán a versengést mérsékelő más szempontból is divergálódhatnak, megváltozhat például az élőhelyük. A poszméhek szívószájszervének hossza például különbözhet a látogatott virágokhoz való alkalmazkodás eredményeképpen, de vannak azonos hosszúságú szívó szájszervvel rendelkező fajok is, ezek rendszerint más-más habitatokat laknak, például különböző magasságokban élnek a hegyvidéken. Brown és Wilson alkotta meg az ötvenes évek közepén a jellegeltolódás (character displacement) kifejezést a földrajzi variációnak egy sajátos mintázatára, amikor két faj szimpatrikus populációi bizonyos jellegekben jobban különböznek, mint allopatrikus populációik. A mintázat kialakulásának egy lehetséges oka, hogy a jelleg valamilyen élelem, vagy más forrás használatával, amiben egyébként a fajok vetélkednének egymással (ugyanezt a kifejezést használják a kompetíció miatt kialakuló divergencia folyamatára is). A galápagosi földi pintyek, a Geospiza fortis és G. fuliginosa jobban kölönbözik csőrének méretében ott, ahol együttesen fordulnak elő, mint ahol külön-külön. A csőr méretének különbözősége korrelál annak hatékonyságával, hogy a madarak hogyan tudják feldolgozni a különböző méretű és keménységű magvakat, és a pintyek populációméretét gyakorta az élelem korlátozza, és fellép a kompetíció 5. ábra.
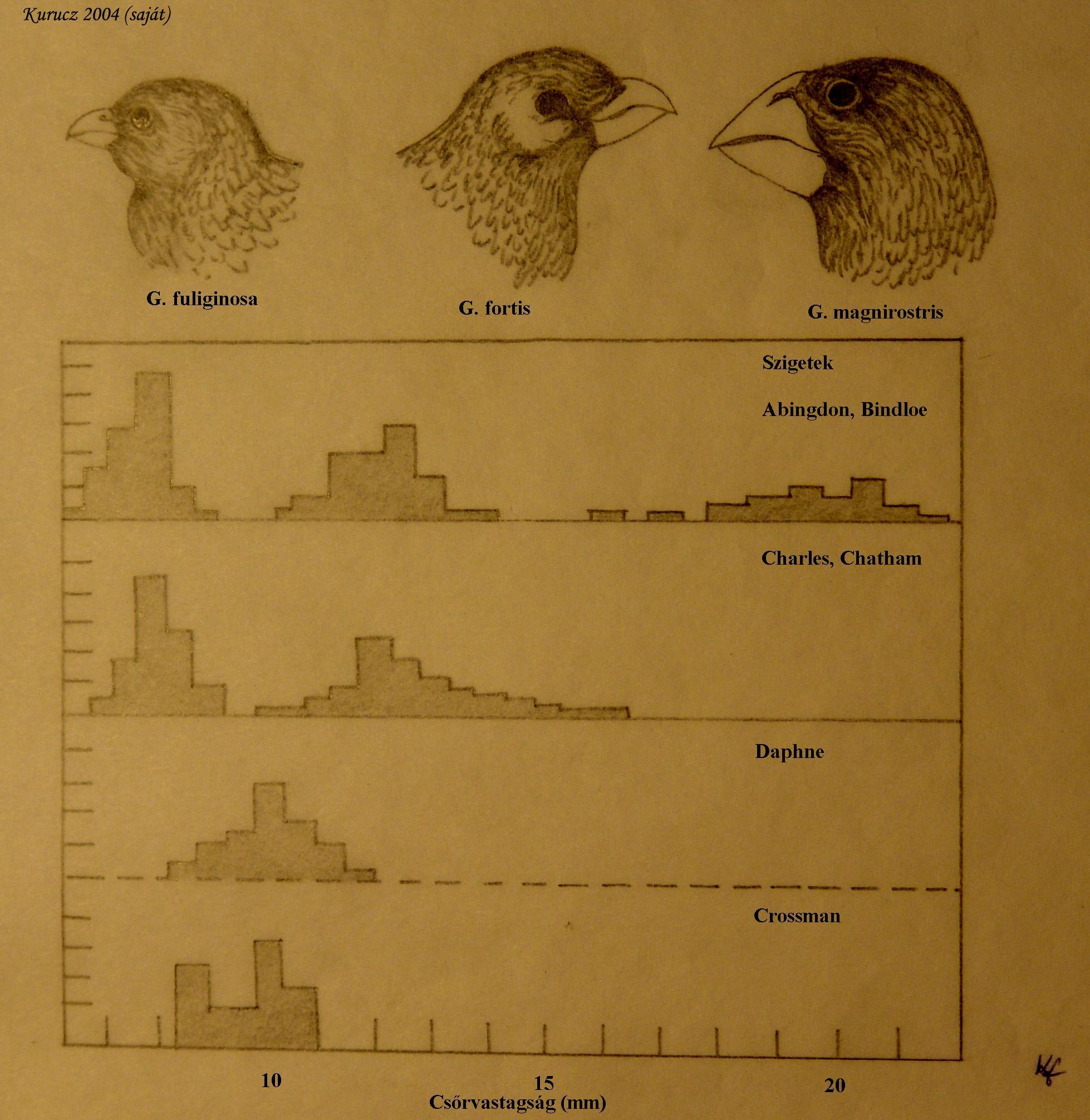
Látványos példa a tüskés pikó (Gastrosteus aculeatus) „komplex” esete. A komplex elnevezés indokolt, mert a faj minden tekintetben szokatlanul változatos, mint az a példából is kiderül. Észak-Amerika északnyugati részén több tóban két, reproduktív izolációban élő forma él: egy bentikus és egy limnetikus. A két forma különbözik a testformában, szájmorfológiában, a kopoltyúfedő szerkezetében. Más tavakban, ahol csak egy forma van jelen, annak morfológiája köztes. Az ökológiai felszabadulás (ecological release) egy másik földrajzi mintázat, amikor egy faj, vagy populáció nagyobb variációt mutat a források felhasználásában és az ehhez társuló jellegekben, ha egyedül van jelen, mintha vele versengő fajok is jelen vannak. A jelenség gyakorta fellép szigeti populációknál. A kókusz-szigeti egyetlen pintyfajnak sokkal szélesebb skálán mozog az étrendje, és többféle módon szerez táplálékot, mint bármely rokon faja a Galápagos-szigeteken, ahol sokkal több faj van. Hasonló módon, a Hispaniola szigetén élő egyetlen harkályfaj nagyobb mérvű szexuális dimorfizmust mutat csőr- és nyelvhosszát illetően, mint a kontinensen együtt élő fajok bármelyike, és a nemek markánsan különböznek abban, hogy hol és mikor táplálkoznak.
6.6 Az ökológusok szerint a versengés formálja leginkább az ökológiai közösségeket, ez a fajok közötti kölcsönhatás magyarázza leginkább a detektálható visszatérő mintázatokat. A versengés általában megakadályozza azoknak a fajoknak az együttes előfordulását, amelyek túlságosan hasonló módon használják fel a forrásokat. Az eredmény a forrásokat következetesen felosztó, meghatározott számú szimpatrikus faj jelenléte. A közeli rokon, nagyon hasonló igényű fajok kölcsönösen kizárják egymás jelenlétét. A versengés feltehetően korlátozza a fajok diverzitását a jelenlegi életközösségekben, és befolyásolhatta a fajok diverzitását evolúciós léptékben is. Az ökológiai kölcsönhatások nagy mértékben irányítják, megjósolható útra terelik a kölcsönhatásban lévő fajok evolúcióját, konvergens mintázatokat alakíthatnak ki. A jelenség tendenciaszerűen lép fel, nem kényszerpályák alakulnak ki: nem alakultak ki: vérszívó denevérek pl. csak Dél-Amerikában élnek, pedig máshol is vannak nagytestű patások is, denevérek is, vagy a sivatagi gyíkok diverzitása drámaian nagyobb Ausztráliában, mint más kontinenseken. Néhány, meglepően következetes mintázat azonban kialakult a konvergens evolúció következtében. A közösség szintjén fellépő konvergencia egyik legszebb példája a nyugat-indiai-szigetvilág anoliszainak példája. Az anoliszok az újvilági trópusokon élő, rovarevő, fán lakó gyíkok. Különböző fajaik versengenek a táplálékért, és a verseny látványosan befolyásolta közösségeik szerkezetét. A Kis-Antillák minden egyes apró szigetén egy, vagy két faj él. Az egyedül élő fajok általában közepes méretűek, míg a nagyobbacska szigeteken egy nagy, és egy kis faj él békésen együtt: különböző méretű és mikrohabitatú rovarfajokat fogyasztanak. A kis méretű fajok közeli rokonok, monofiletikusak, és a nagy termetűek szintén monofiletikusak egymás között, így látható, hogy két klád adott egy-egy fajt minden szigetnek. A Nagy-Antillák nagy szigetei (Kuba, Hispaniola, Jamaika, Puerto Rico) sok fajnak adnak otthont. Az anoliszok bizonyos mikrohabitatokat foglalnak el: fakorona, ágak, törzs. A mikrohabitatok fajai minden szigeten eltérnek. A különböző mikrohabitatok lakóit ökomorfoknak (ecomorphs) nevezzük, következetes adaptív morfológia jellemzi őket. Az ökomorfok ismételten kifejlődtek, az egyes szigetek különböző ökomorfjai monofiletikus csoportot alkotnak, tökéletes párhuzamban más szigetek, más monofiletikus csoportot alkotó fajaival. A mintázat legkézenfekvőbb magyarázata, hogy a szigetekre kerülő új fajok hasonló evolúción estek át, hasonlóan adaptálódtak az előzőleg kihasználatlan mikrohabitatokhoz, hogy elkerüljék a kompetíciót. Hasonló az Accipiter-fajok esete, amelyeknek testmérete erősen különböző, ahogy áldozataik testmérete is. Szimpatrikus fajaik mérete mindig jelentősen különbözik. Az ilyen és hasonló példák alapján az ökológiai elvek figyelembevételével az evolúció menete sokszor megjósolható.
6.7 Több fajt érintő kölcsönhatások (Multispecies interactions) J. Thompson (1999) figyelte meg, hogy egy bizonyos fajra más fajokkal való interakciói által kifejtett szelekciós nyomás változhat egyik földrajzi populációról a másikra, az eredmény a koevolúció egy földrajzi mozaikja. A szelekció lehet erőteljesebb bizonyos helyeken, mint máshol, esetleg más jellegeket részesíthet előnyben, sőt, a génáramlás a különböző módon alkalmazkodott populációk között lokálisan nem megfelelő adaptációk megjelenéséhez is vezethet. Benkman és munkatársai kölcsönhatásoknak egy ilyen földrajzi mozaikját vizsgálták. A szereplők: Pinus contorta fenyő, és két magevő, egy észak amerikai vörös mókus (Tamiasciurus hudsonicus) és a keresztcsőrű (Loxia curvirostra). Az északi Sziklás-hegységben, a fenyő elterjedési területének döntő részén, a mókusok szüretelik le és tárolják a tobozok döntő részét, és ők a magvak elsődleges fogyasztói. A mókusok a keskenyebb tobozokat kedvelik, amelyeknél a magvak tömegaránya magasabb, így tkp. A szélesebb, kevesebb magvú tobozokat juttatják szelekciós előnyhöz. A csaknem kizárólag fenyőmaggal táplálkozó, erre szakosodott csőrrel rendelkező keresztcsőrű sokkal gyakoribb azokban a hegyvonulatokban, ahol a mókusok hiányoznak. A madarak kevésbé hatékonyan táplálkoznak a nagyobb, szélesebb tobozokból, amelyeknek vastagabbak a pikkelyei — és azok a fenyőpopulációk, amelyeket csak a keresztcsőrűek dézsmálnak, pontosan ilyen tobozokkal rendelkeznek. Az itteni keresztcsőrűek csőre hosszabb és mélyebb, mint azokon a részeken, ahol mókusok is vannak. A hosszú, mély csőr a táplálkozási teszteknél is hatékonynak bizonyult a nagyobb, vastag pikkelyes tobozoknál. A fenyők és a keresztcsőrűek koevolúciója tehát nyilvánvaló, ahol ezek kölcsönhatása erőteljes, de ahol a mókusok a dominánsak, ott ők irányítják a tobozok morfológiai jellegeinek változását, és a keresztcsőrűek aszerint alkalmazkodnak 6. ábra
8.6. ábra - Két faj, amelyek befolyásolják a Pinus contorta tobozainak morfológiáját: keresztcsőrű és kanadai vörös mókus
 |
 |
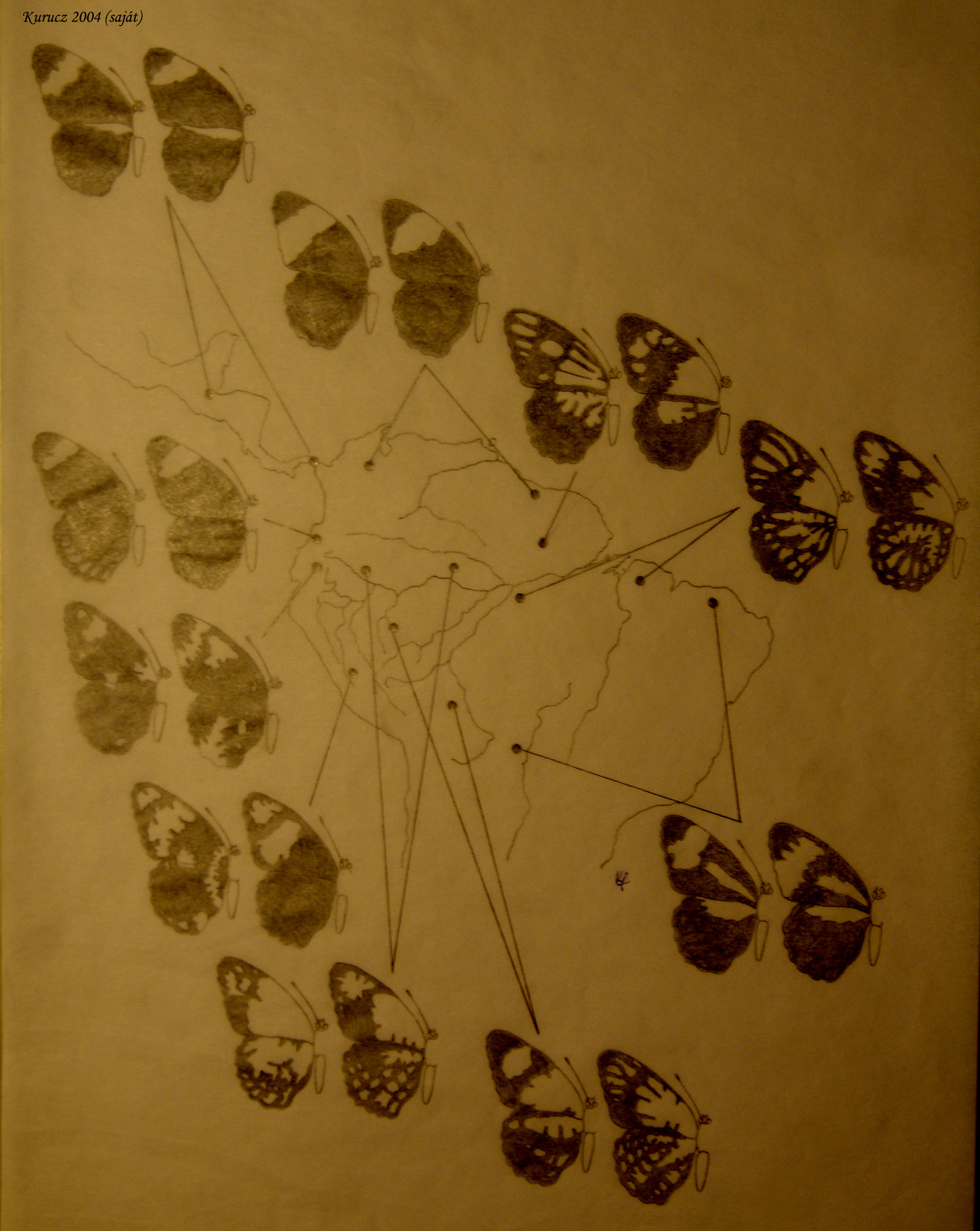
A defenzív mimikrinél egy vagy több faj nyer védelmet a ragadozóktól azáltal, hogy hasonlít egy másik fajra. Hagyományosan két formáját különítik el, a Bates-i („batesiánus”) mimikrinél egy ehető faj hasonlít egy ehetetlen fajra (előbbi a „mimic”, vagy mímelő, utóbbi a modell. A mülleri mimikrinél két, vagy több ehetetlen faj alkot ko-modellt (vagy „co-mimic”-et). Mindkét esetben kellemetlen tapasztalataikból a ragadozók megtanulják a kellemetlen ízű fajra emlékeztető potenciális préda elkerülését. A modellek és mímelők gyakorta viselnek figyelemfelkeltő, figyelmeztető jeleket (aposzematikus mintázatok). A védekező mimikriből hasznot húzó fajok csoportjai „mimikri gyűrűk” (mimicry rings). Sok olyan eset ismert, amikor a mimikri gyűrűk magukba foglalnak erősen, és kevésbe rossz ízű fajokat, utóbbit nevezhetjük előbbi „kvázi batesiánus” mímelőjének. Sokszor több mimikri-gyűrű is jelen van ugyanabban a régióban, mindegyiknek több, hasonló mülleri mimikrit viselő tagja van, és gyakorta foglalnak magukba ehető batesiánus mímelőt is (vagy enyhén rossz ízű kvázi-batesiánus mímelőt). Különösen a fecskefarkú lepkéknél a batesiánus mímelők néha polimorfak a populációban, minden egyes morf egy másik ehetetlen fajra hasonlít. A mülleri mímelők szinte soha nem polimorfak a populáción belül, de egyes fajok földrajzi alfajainak más riasztó jelzései lehetnek, és más-más mimikri gyűrűhöz tartozhatnak. A mimetikus fenotípusra ható szelekció mértéke függ a mímelő gyakoriságától a modellhez viszonyítva, és a modell rossz ízének, fogyaszthatatlanságának mértékétől. A gyakoribb ehetetlen modellel szerzett újabb tapasztalat nyilván riasztó. Ugyanakkor ha a modellre hasonlító jóízű mímelővel volt utoljára pozitív élménye, aligha kerüli el a következő mímelőt. Ha ritka az ehető mímelő, akkor kisebb az esélye, hogy pozitív megerősítést ad, így előnyösebb az ehetetlen modellhez hasonlítania. (A rossz íz mértéke is befolyásolja a fejleményeket.) A mimetikus polimorfizmus tehát gyakoriság-függő szelekcióhoz vezet. Miután a mülleri mímelők felerősítik a ragadozók averziós tanulási folyamatát, így valószínűleg stabilizáló szelekció érvényesül minden szimpatrikus, ehetetlen faj mintázatára. Sok, látszólag mülleri mímelő azonban tulajdonképpen épphogy rossz ízű, kvázi-batesiánus, így rontja a figyelmeztető jelzések hatásfokát. Az ilyen, kvázibatesiánus fajok mutáns fenotípusai előnyösek lehetnek, ha egy másik mimikrigyűrű még gyakoribb, vagy még ehetetlenebb fajára hasonlítanak 7. ábra.
8.7. ábra - A Heliconius nem két faja párhuzamos „mimikai gyűrűt” formál Dél- és Közép-Amerikában. A megjelölt helyeken mind a H. erato, mind a H. melpomene jelen van, és egymáshoz hasonul, vagyis a különböző helyeken a két faj párhuzamosan változik. Mindkét faj mérgező, így a Müller-féle mimikri gyönyörű példájával állunk szemben
 |
A kakukk gazdafajai általában nem kezelik az övéiktől különbözően a parazita fiókáit, de némelyik gazdafaj felismeri a parazita tojásait, és vagy kilöki őket vagy otthagyja fészkét és teljesen új fészkelésbe kezd. A fészekparazitás leglátványosabb ellenadaptációja a tojásmimikri. Az európai kakukk (Cuculus canorus) minden egyes populációjában különböző genotípusú egyedek vannak, amelyek mindegyike más-más gazdát részesít előnyben. Ezek a gazdafajéhoz hasonló tojásokat raknak. Más egyedek tojásai viszont nem mutatnak hasonlóságot semmilyen gazdáéval. A mesterséges kakukktojások sorsának nyomon követésével Davies és Brook (1998) azt találta, hogy azok a fajok, amelyeket a kakukk egyáltalán nem használ gazdaként (nem fér hozzá a fészekhez stb.), általában nem lökik ki a tojásokat, míg a kakukk kedvelt gazdafajai közül azok, amelyeknek tojását a kakukk utánozza, gyakrabban lökik ki a mesterséges tojásokat. Ezek a fajok nyilvánvalóan alkalmazkodtak a fészekparazitizmushoz, mi több, Izlandon, ahol kakukkok egyáltalán nem élnek, a gazdafajok elfogadták a mesterséges tojásokat, míg Nagy-Britanniában, ahol ugyanezek a fajok fontos gazdák, elutasították a mesterséges tojásokat. Érdekes módon a gazdaként gyakran és ritkán használt fajok között nem volt különbség a tojások megkülönböztetését tekintve. Valószínűnek tartják, hogy a ritkábban parazitált faj nyerésre áll a kakukk elleni evolúciós harcban: régebben népszerűbb volt a kakukkok körében, de a tojások felismerésének képessége túlságosan nagy rizikót jelent a parazita számára.