Tartalom
Darwin szerint az evolúciós változás a különböző életformáknál folyamatos, de a változás sebességéről nem voltak ismeretei. Azt mindenesetre sohasem állította, hogy minden vonalban ugyanolyan rátával zajlik az evolúciós változás, vagy a földi élet evolúciója során minden periódusban egységes lenne az üteme. A vonalak morfológiai változásának és diverzifikálódásának üteme drámaian különbözhet. Míg a bojtosúszós halak – az indonéziai és afrikai Coelocanthák összevetése alapján – az utóbbi 5-10 millió év alatt alatt gyakorlatilag nem változtak (igaz, a színük különböző) (1. ábra) addig a hawaii-i gyapjasmadaraknak (2.ábra) hasonló időintervallumban 50 különböző színű, formájú, és életmódú faja alakult ki, .
7.1. ábra - Az afrikai bojtosúszós hal (Latimeria chalumnae) és az indonéziai bojtosúszós hal (L. menadoensis)
 |
 |

George Gaylord Simpson (1902-1984) paleontológus sok csoportot vizsgált, és elbűvölte az a drámai különbség, ami különböző biológiai csoportok evolúciós rátájában tapasztalható. Simpson megalkotta a „tempo and mode of evolution” („az evolúció sebessége és módja”) kifejezést, ami kifejezi, hogy az evolúciós változás sebessége és típusa nem azonos a különböző vonalakban az evolúciójuk különböző szakaszaiban, vagy különböző élőhelyeken. Simpsont érdekelte, hogyan lehet a paleontológia és genetika együttes alkalmazásával többet megtudni az evolúciós változás rátájáról és mechanizmusairól.
A különböző vonalak, periódusok vagy helyek jellemző evolúciós tempójának és módjának jellemzéséhez meg kell becsülni az evolúciós változás rátáját. Miután a(z evolúciós) ráta tulajdonképpen bizonyos távolság és az eltelt idő hányadosa, az evolúció rátájának becsléséhez szükség van a felgyülemlett evolúciós változások mérésére és annak az időnek az ismeretére, ami alatt a változások történtek. Simpson időskálája abból következett, hogy fosszilis anyag vizsgálatából vonta le következtetéseit, így olyan taxonokat vetett össze, amelyekről földtörténeti mércével hosszú időt felölelő ideig folyamatos fosszilis anyag állt rendelkezésére. Kimutatta például, hogy a lovak fogainak dimenziói bizonyos időszakokban gyors tempóban változtak, míg máskor lassabban, és megmutatta, hogy a lovak fajképződése (speciációja) többszörös sebességgel zajlott az ammoniteszekéhez képest.
A fosszilis anyag azonban nem ad teljes és folyamatos képet az evolúció történetéről. Szerencsére a genom az evolúciós történet egy alternatív, kiegészítő, a paleontológiai anyaggal összevethető forrása. A DNS-szekvenciák összevetésével, bizonyos változások egyenletes ütemű megjelenését feltételezve, azaz a szekvenciákat molekuláris órákként alkalmazva, az evolúció tempója és módja vizsgálható, ha a genomi változások rátáját ismert evolúciós időskálára vonatkoztatjuk. Mivel a fosszilis anyag a különböző taxonok megjelenésének egyetlen megbízható forrása, a jól azonosítható, esetleg átmeneti formákat reprezentáló kövületek használhatók az időbeli viszonyok kalibrálására.
Minden genom hordozza evolúciós történetének emlékeit. A DNS szekvenciák módot adnak arra is, hogy az evolúciós események idődimenzióit megbecsüljük. Mivel a molekuláris változások folyamatosan gyűlnek fel,azonos szekvenciákat alapul véve feltehetően azonos rátával, megjósolható, hogy két olyan vonal genomja, amelynek időben közeli közös őse van, kisebb mértékben különbözik, mint a távolabbi rokon vonalak. Pontosabban: ha meg tudjuk becsülni, hány genetikai változás történt, mióta a két vonal elvált, és tudjuk, milyen gyorsan akkumulálódtak a változások a vonalakban (esetleg több, meghatározott szekvenciát alapul véve), a genetikai változás mennyisége felhasználható a két vonal elválási idejének becslésére.Ezen az elven alapul a filogenetikus fák előállítása, ami egy külön jegyzet tárgyát képezi, és csak röviden szólunk róla a fejezet végén.
Az evolúciós időhossz molekuláris adatokból való kikövetkeztetése mindig is ellentmondásos téma volt: a „molekuláris órákat” sokan pontosnak, sokan pontatlannak tartják.
A fejezetben majd röviden foglalkozunk a kambriumi robbanással, mint a földi élet szempontjából meghatározó eseménnyel
Az alapvető kérdések közé tartoznak, hogy el lehet-e számolni a „robbanásokkor” lezajló változások sebességével a mai evolúciós rátákkal, valamint hogy miért fejlődtek egyes vonalak gyorsabban?
A fosszilis bizonyítékok tehát felhasználhatók bizonyos vonalak formai és méretbeli változásainak becslésére.
Simpson az ötvenes években kezdte meg az evolúciós ráták tanulmányozását. Különbséget tett az egyes sajátságok, vagy sajátság-együttesek fejlődési rátái között, -- ezeket filogenetikai rátáknak (phylogenetic rates) nevezte el --, és a taxonómaia ráták (taxonomic rates) között. Taxonómiai rátákkal jellemezhető a különböző jellegekkel rendelkező taxonok kialakulása, kihalása, és másokkal való helyettesítődése.
Az egyedi jellegek evolúciós változásainak rátája nagyban változó a különböző jellegek, vonalak esetén, sőt, ugyanabban a vonalban is az idők folyamán.
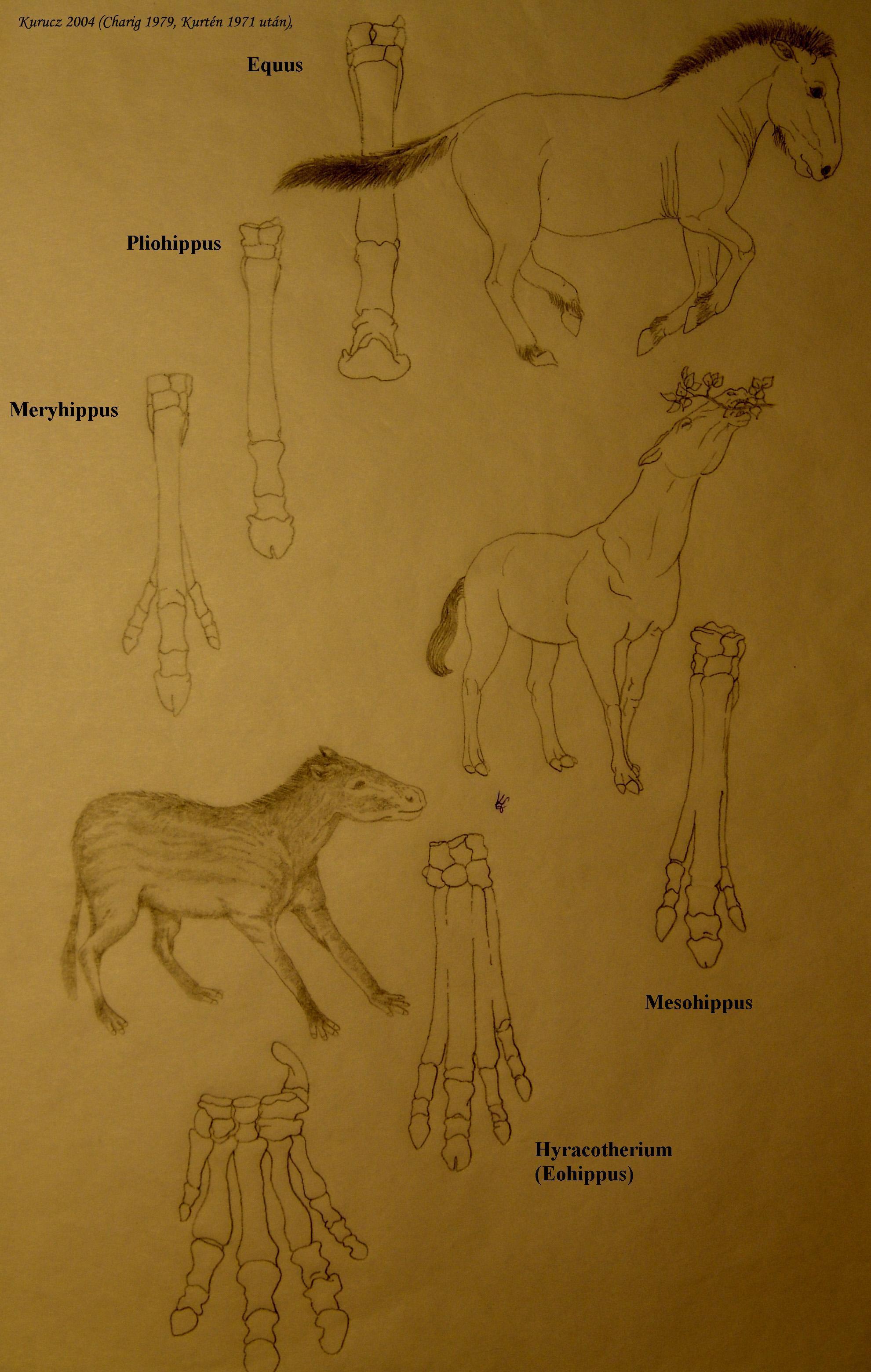
Lovak családjának 6-8 modern faja van és leszármazási vonalaik jól ismertek
Szép példa az Equus → Merychippus→Mesohippus→ Hyracotherium (utóbbi 55 millió éve élt) vonal 3. ábra. Az egyes lófajok fogai jellemző, meglehetősen tagolt morfológiájúak, külön előnyük, hogy fosszilis fogak szép számban kerültek elő.
7.3. ábra - Lovak és lábaik alakulása: A Hyracotherium (alul) az első igazi lóféle kutya méretű volt. Lábain négy ujj nőtt, amelyek közül egy anatómiai szempontból a harmadik kissé hosszabb volt, mivel növekedési rátája meghaladta a többiét. Ahogy a lófélék mérete nőtt, az oldalsó ujjak elvesztették kapcsolatukat a talajjal, a harmadik ujj metakarpális ízének allometrikus növekedése miatt. Később az oldalsó ujjak elcsökevényesedtek egy további genetikai változás miatt.
 |
Az eocénben a lovak maximum kutya méretűek, minimum macskányiak voltak - fogaik is kisebbek és mások, mint kései utódaiké. Mivel a leszármazási viszonyok ismertek, az evolúciós változás direkt méréssel megadható. MacFadden klasszikus tanulmányaiban 408 fogat vizsgált, 26 (jó okkal feltételezett) ős–leszármazott párból.
X1 korai minta átlaga
X2 későbbi minta átlaga
r = (ln X2 – ln X1)/Δt
A ráta pozitív, ha a jelleg evolúciósan „nő”, negatív, ha „csökken”.
Sokszor csak az abszolút érték számít.
Haldene javaslatára az : „darwin” az evolúciós változás mértéke:
1 darwin egység a jelleg e » 2,718-szoros (a természetes logaritmus alapszámának megfelelő) változása 1 millió év alatt. (A gyakorlat számára alkalmazható az 1 darwin= 0.1 % változás a jelleg értékében 1000 év alatt definíció.).
pl. X1 = 1; X2 = 2,718; t= 10 millió év
r = (ln 2,718 – ln 1)/10 (millió év)= 0,1 darwin
Ha nem a logaritmusokat vennénk, a jelleg evolúciója gyorsabbnak tűnne. Pl. ha egy méret abszolút értékben nagyobb lesz egy eleve nagyobb termetű fajokat felvonultató sorozatban, akkor a kisebb termetűekből álló sorozathoz képest nagyobbnak tűnik a változás, akkor is ha arányaiban ugyanolyan a változás mértéke (ha mindegyik vonalban másfélszeresére változik, az az eleve nagyobbnál abszolút értékben nagyobb változást jelenthet). A logaritmikusan transzformált méretek figyelembevételével különböző méretű állatok is összevethetők.
0,05 – 0,1 darwin általában 15-30%-nyi változásnak felel meg 1 millió évente.
A darwinnak is vannak hátrányai, így a kutatók újabban az evolúciós változást a generációnként mérhető standard deviációk számának változásával mérik, ezt az egységet Gingerich után Haldane-nek nevezték el.
Amikor az evolúciós rátákat definiált korú, ős-leszármazott fajpárok között mérték, az első tapasztalat az volt, hogy az evolúció átlagos rátája igen alacsony. Pl. a korai eocénben élő ló, a Hyracotherium grangeri első őrlőfogának mérete fluktuált, de 650 000 év alatt alig változott, átlagos változása csak 0.0000057 haldane volt 113 000 generáció átlagában, majd 0.0003 haldane 6745 generáció átlagában. A fentiek nagyon kis értékek, de általánosak a kövületek körében. A hosszú periódusok azonban elrejthetnek rövid idő alatt zajló fluktuációkat. A rövid intervallumokon mért értékek magasabbak: két Hyracotherium faj 13500 generációs átmeneténél mért 0,00024 haldane érték magasabb, minta a H. grangerinél mért legmagasabb érték.
A fosszilis anyag változásai tehát nagyon lassúak. A hosszú intervallumokon mért ráták múlhatatlanul alacsonyabbak, mint az ember által bolygatott, áttelepített, évszázados skálán mérhető változások. Az egyik leggyorsabb evolúciós rátát a házi verebek csontvázain mérték, amelyeket az USA-ba Európából vitték be a XIX. században. Szárny- és csőrméretük legfeljebb 0,024 haldane növekedést mutatott. Rajtuk is túltettek azok a bodobácsok (soapberry bug) 4. ábra, amelyek birtokba véve az új, betelepített növényeket 0,01- 0,035 haldane-os növekedést mutattak, ami százszorosa a Hyracotheriumok leggyorsabb ütemű fognövekedésének. Ez a ráta is sokkal lassúbb, mint ami laboratóriumban elérhető. A fosszilis emlősöknél mért ráta huszada annak, mint amit a verebeknél mértek, a dinoszauruszok növekedési rátája pedig tízszer kisebb.

Ha a jellegek, még akár jóval lassabban, mint az itt említettek bármelyike, egy irányba növekednének, az élőlények még a mostaninál is jobban különböznének, az egér mérete meghaladhatná a legnagyobb dinoszauruszét is. Az evolúciós változások igen gyorsak is lehetnek, de a magas ráták a tapasztalatok szerint sohasem maradnak fenn túl hosszú ideig.
Evolúciós ráták változóak: braditél – lassú; horotél - tipikus (lovak); tachitél – gyors.