A fosszíliákkal foglalkozó őslénytan 1822-ben különült el a „szedimentekkel”, azaz a bezáró kőzetekkel foglalkozó geológiától. Nevét (paleontológia) 1831-ben Brongniarttól kapta, de a tudományos őslénytan megalapozója valójában Cuvier (1759–1832) volt. Az őslénytan – tárgyát tekintve – a biológiához áll közel, alkalmazási területei azonban a geológiához kötik.
Az őslénytan legfontosabb alkalmazási területe a biosztratigráfia, vagyis relatív kormeghatározás és az őskörnyezettan. Évszázados tudományos tapasztalatok, sziszifuszi munka eredményeit hasznosítjuk. A látványos ősmaradványok ritkák, de van néhány világhírű előfordulás.
A 600-550 millió éves, vendi korszakbéli Ediacara-hegység kövületei (129. ábra) lágytestűek voltak, ezért különleges körülmények kellettek a fosszilizációhoz. Rendszertani besorolásuk nehézségekbe ütközik, mert látszólag nincsenek kapcsolatban a mai élővilággal. Legérdekesebb, hogy testnyílás nem található rajtuk, ezért egyesek azt feltételezik, hogy fotószintetizáló lényekkel éltek belső szimbiózisban.
 |
129. ábra Az ediacara-fauna rekonstrukciója
130. ábra. Hallucigenia, az ediacara-fauna képviselője.
Dél-Németország bővelkedik csodálatos jura időszaki fosszíliákban. Sok példánya került elő a tengeri ragadozó hüllő Ichthyosaurusnak (131. ábra).
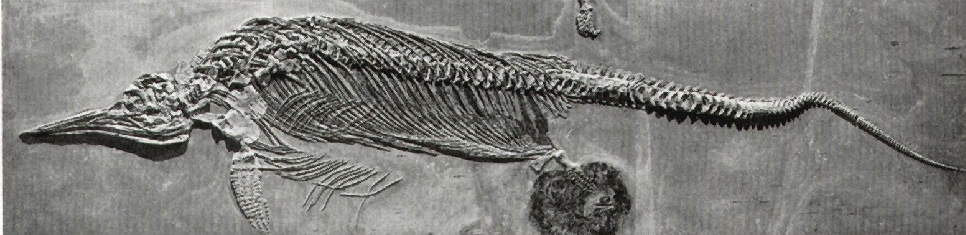 |
131. ábra. Tengeri ragadozó hüllő (Ichthyosaurus) maradványa az alsó-jura holzmadeni posidoniás palából
131. ábra. Tengeri ragadozó hüllő (Ichthyosaurus) maradványa az alsó-jura holzmadeni posidoniás palából
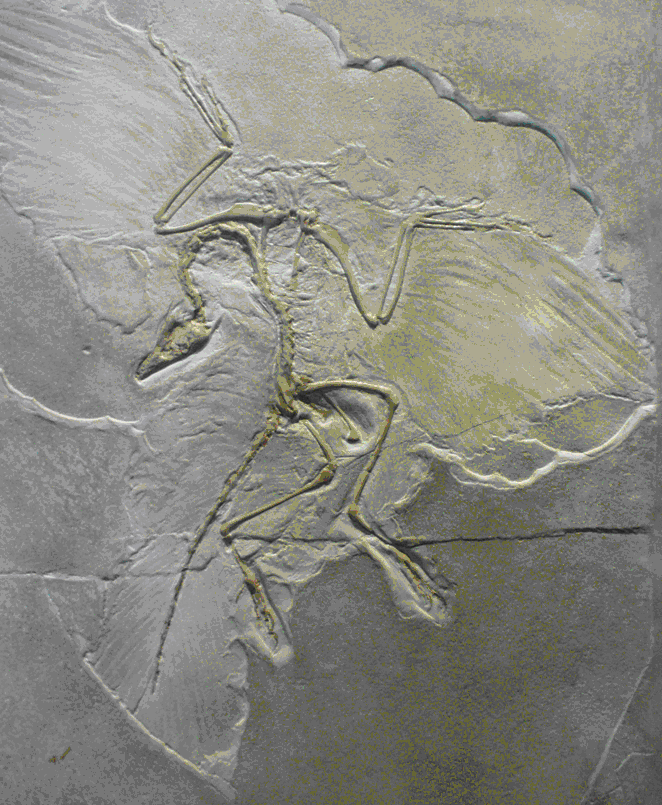
A bajorországi Solnhofenben viszonylag ritkák a kövületek, viszont mindegyik rendkívül jó állapotú. Leghíresebb és a tudomány számára legfontosabb lelete az Archaeopteryx, az ősmadár (132. ábra). Első példányát néhány évvel a darwini evolúcióelmélet megjelenése után találták, annak fényes bizonyítékául szolgált. Az Archaeopteryx a mozaikevolúció folyamatát példázza: felépítésében hüllő- és madárbélyegek egyaránt előfordulnak.
Eocén emlősök csodálatos épségen megőrződött maradványait ismerjük a németországi messeli palából (133. ábra).
 |
133. ábra. Eocén hangyász (80 cm) és sünszerű rovarevő (40 cm) rekonstrukciója a jobb oldalon látható, rendkívül jó megtartású messeli fosszíliák alapján
Magyarországon is találunk olyan lelőhelyeket, amelyek nemzetközi érdeklődésre is számot tartanak. Ipolytarnócon miocén vulkáni tufaszórás üledékei őrizték meg egy tóparti élővilág nyomait. A 4,2 millió éves Pulai Alginit nevű kőzetben – az egykori vulkáni krátertavakban leülepedett, főként algavázakból álló képződményből előkerült egy kitűnő megtartású orrszarvú fosszília, amelyet a Zirci Múzeumban őriznek. Világhírre tett szert a bükkábrányi lignitbányászat során feltárult mocsárerdő maradványa (151. ábra).
Az évszázados kutatások során megismert számtalan ősmaradványt rendszerezni kellett. A biológiai rendszerezés alapja sokáig tipológiai volt, azaz a megállapított közös vonás, tulajdonság, típus. A ma használatos filogenetikai osztályozás a tényleges rokonsági kapcsolatokra épül. Ennek megállapítása a fosszíliák körében – elsősorban a lágytest hiánya miatt – gyakran nagy nehézségekbe ütközik. Vannak olyan maradványok, amelyek rendszertani besorolását még törzsi szinten sem tudjuk elvégezni!
Az osztályozás elvével és módszereivel a taxonómia foglalkozik. A rendszertan hierarchikus rendben kategóriákat (rendszertani egységeket), azokon belül taxonokat (csoportokat) különít el (9. táblázat)
14.1. táblázat - 9. táblázat. Az élővilág rendszerezésének (szisztematika) kategóriái, példákkal
|
Regnum (birodalom) |
állatok |
|
Phylum (törzs) |
Mollusca (puhatestűek) |
|
Classis (osztály) |
Cephalopoda (fejlábúak) |
|
Subclassis (alosztály) |
Ammonoidea (ammoniteszek) |
|
Ordo (rend) |
Phylloceratida |
|
Familia (család) |
Phylloceras-félék |
|
Genus (nem, nemzetség) |
Phylloceras |
|
Species (faj) |
Ph. heterophyllum |
Névadás, nevezéktan (nomenclatura) és rövidítések
Az érvényes nevezéktan Linné Systema Naturae című munkájának X. kiadásával (1758) kezdődik. Az előtte keletkezett nevek nem használhatók. A nevezéktan kétnevű (binominális nómenklatúra). Minden állatfaj neve a nagybetűvel írott nemzetség (genus) és kisbetűvel írott faj (species) nevéből áll. Teljes névhasználatban ezt követi a leíró neve és a leírás évszáma: Balatonites balatonicus (Mojsisovics 1873).
A következő oldalakon a geológiai gyakorlatban fontos csoportok közül mutatunk be néhányat – azt szemléltetendő, milyen lehetőségek adódnak a szilárd váz alapján a rendszerezésre, milyen következtetéseket tudunk levonni a váz felépítéséből.
Szilárd, kamrázott, nyílásokkal áttört váz. Általában mikroszkopikus méretűek, de az óriásforaminiferák közül a Nummulites a 10 centiméteres átmérőt is elérheti.
Jellemzően normál sótartalmú, meleg, sekélytengeri bentosz lények, a planktoniak nyílttengeriek.
Első megjelenésük a kambriumban volt.
Minthogy a foraminiferák általában szűktűrésű szervezetek, előfordulásukból következtetni lehet az egykori tengeri környezetre: az aljzat jellegére, az üledékképződésre, a hőmérsékletre vagy akár a vízmélységre.
 |
134. ábra. Különböző „kisfora¬miniferák”,a vázak mérete kb. 0,5-1 mm
A korallok egyedei (korallit) külső mészvázat választanak el, amely a hengerszerű külső falból (téka), és az azt erősítő sugárirányú belső válaszfalakból (szeptum) áll. A több egyedből álló telep a korallum, változatos alakja függ a környezettől.
Szűktűrésű szervezetek, ezért kiváló környezetjelzők: jellemzően meleg, normál sótartalmú, oxigéndús, sekély tengervízben éltek.
A kagylók kétoldali részarányos felépítésűek, két teknőből álló vázuk van, amelyek a köpenyüreggel együtt a lágytestet védik.
A kagylók többsége fenéklakó. Az édesvizektől a túlsós tengerekig, a parti vizektől az abisszikus régióig mindenhol előfordulnak, a szárazföldet azonban nem tudták meghódítani.
A kagylók a kambrium elején jelentek meg, virágkoruk a mezozoikumban volt. Jelentős kőzetalkotók, jó környezetjelzők.
A csigák egy részből álló, spirálisan felcsavart, kamrázatlan vázat építenek. A váz anyaga általában aragonit.
Többségük sekélytengerekben él (epibentosz vagy inbentosz), de alkalmazkodtak az édesvízi és (a puhatestűek közül egyedüliként) a szárazföldi élőhelyekhez is.
A csigák gyakran fordulnak elő kőzetalkotó mennyiségben. Jól jelzik az egykori környezetet, a sótartalmat, hőmérsékleti viszonyokat.
A fejlábúak a puhatestűek legfejlettebb csoportját alkotják. Földtani szempontból a legfontosabb alosztályuk az ammoniteszek. Aragonit anyagú vázuk három részből áll: embrionális kamra, szeptumokkal tagolt kamrázott vázrész és lakókamra (135. ábra, 136. ábra). A szeptumok által lezárt kamrák gázzal voltak telítettek, ami elősegítette a lebegést. A váz többnyire egy síkban felcsavaródó. A kamraválaszfal felépítése a törzsfejlődés során egyre bonyolultabbá vált (137. ábra).
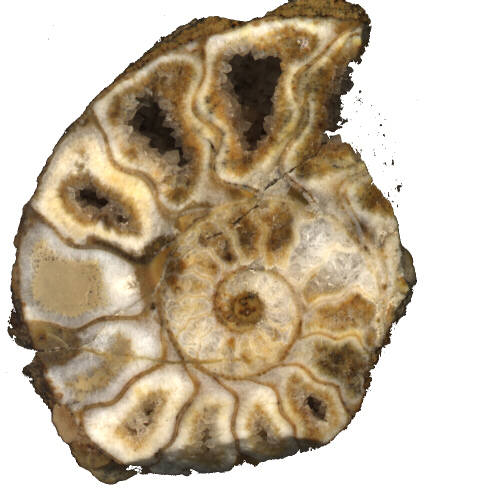 |
135. ábra. Ammonites (Villány, Villányi Formáció) metszete. A kamrákat a kőzettéválás során részben vagy egészében kalcit töltötte ki
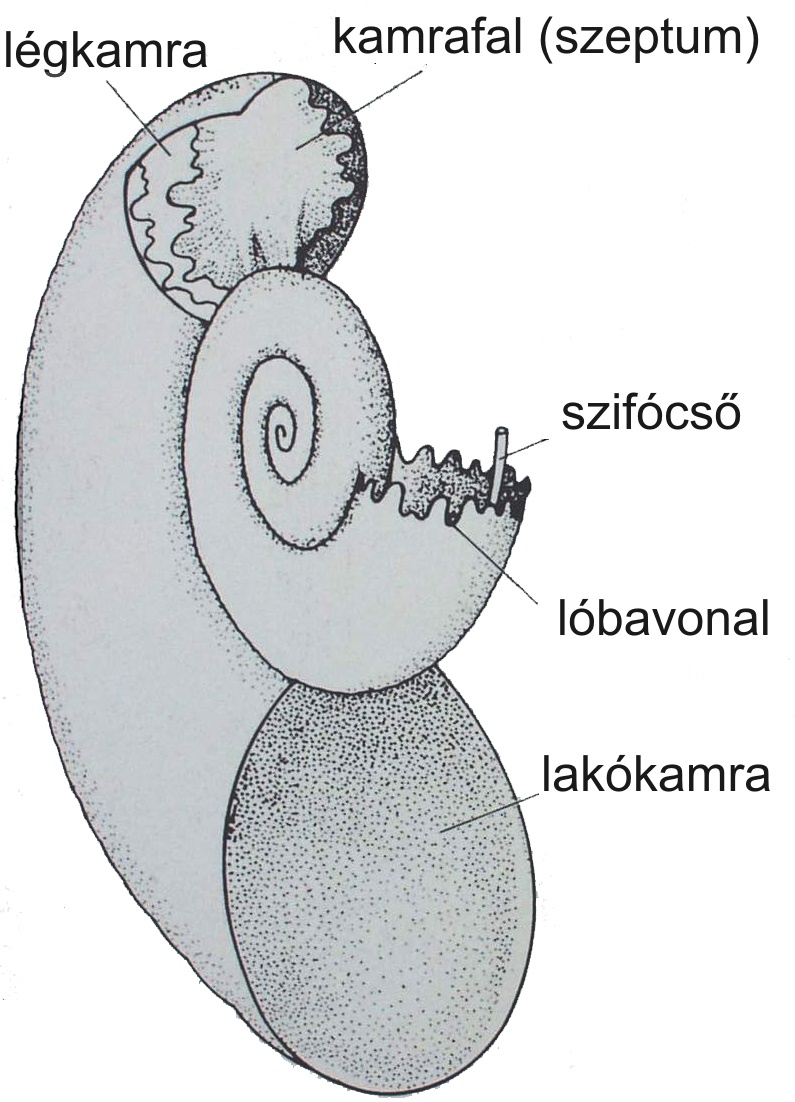 |
136. ábra. Az ammoniteszváz felépítése
Az ammoniteszek úszó életmódot folytattak, a nyílt tengereket népesítették be. A kora devonban jelentek meg, virágkoruk a jura időszakban volt. A kréta végén kihaltak.
Az ammoniteszeket – gyors evolúciójuk és széles körű elterjedésük alapján – a késő-paleozoikum és a mezozoikum biosztratigráfiai tagolására használják. Kitűnő szintjelzők, egy ammoniteszre alapozott zóna százezer év nagyságrendű időtartamot képvisel.
 |
137. ábra. A lóbavonalak (kamra-válaszfalak) lefutásának bonyolultabbá válása az evolúció során. Balra devon Goniatites, középen triász Ceratites, jobbra jura Holcophylloceras lóbavonalai láthatók természetes nagyságban
Az ízeltlábúak az állatvilág jelenlegi leggazdagabb törzsét alkotják. Általános jellemzőjük a szelvényezett test, a hármas testfelépítés (fej, tor, potroh), az ízelt láb és a kitinváz. Négy altörzsük közül a trilobiták és a rákfélékhez tartozó kagylósrákok a legfontosabbak földtani-őslénytani szempontból.
Háromkaréjos ősrákok altörzse (Trilobita)
Elliptikus vagy tojásdad alakú, szelvényezett test. A váz hosszanti irányban is három részre tagolódott (138. ábra). Nagyságuk 3 és 70 cm között változott.
A trilobiták kizáróag tengeriek, a többségük az aljzaton mozgott.
A trilobiták a kambriumban (esetleg már a prekambrium végén?) jelentek meg. Virágkoruk a késő-kambriumban és az ordovíciumban volt, fokozatos hanyatlásuk a szilurtól a perm végi kihalásukig tartott.
A trilobiták a kambrium legjobb szintjelzői, emellett ősföldrajzi (paleobiogeoráfiai) elemzésekre is alkalmasak.
 |
138. ábra. Ordovíciumi Asaphus trilobites kb. kétszeres nagyításban
139. ábra. A mai madárpókok fejlődése,
A brachiopodák váza két teknőből áll, amelynek anyaga az ősi típusok esetében kitinfoszfát, a fejlettebbeknél mész. A teknők legidősebb része a búb. A nyél kilépésének a helye a két teknő között, vagy a háti teknőn található (140. ábra).
A brachiopodák többségére a tengerfenéken, nyéllel (egyes formáknál cementációval) rögzített életmód jellemző. Többnyire sziklára, szilárd aljzatra települnek, de előfordulnak lágy aljzaton is. Általában sekélytengeriek.
A kambrium elején jelentek meg, virágkoruk a paleozoikumban volt.
A brachiopodák jelentős kőzetalkotók és környezetjelzők, a paleozoikumban korjelzők is.
 |
140. ábra. Pygope a bakonyi kréta időszaki képződményekből. A kép szélessége kb. 2 cm. Jól látható a búbon a brachiopodákra jellemző lyuk (foramen), ami a nyélkilépés helye
A tüskésbőrűek jellemzője a vízedényrendszer (ambulakrális rendszer), a kalcitlemezekből álló, ötsugaras felépítésű belső (!) váz és a tengeri, bentosz életmód. A vázelemeket kalcit egykristályok alkotják, amelyek a növekedés során kalcittűk összeolvadásával keletkeznek. Egyes egyedek vázelemeinek a száma a kétmilliót is elérheti. Legfontosabb két osztályuk a tengeri liliomok (Crinoidea) és a tengeri sünök (Echinoidea).
Tengeri liliomok osztálya (Crinoidea)
A crinoideák kalcit anyagú belső váza három részből áll: kehely, a kehely alsó részéhez illeszkedő nyél és gyökér (141. ábra).
A crinoideák szilárd aljzaton, csendes vizű tengeri lagúnák, oxigénben gazdag környezetben, rögzült életmódot folytatnak.
A tengeri liliomok az ordovíciumban jelentek meg, virágkoruk a karbon volt.
A krinoideás mészkövek egyértelműen tengeri környezetre és sekély vízmélységre utalnak.
 |
141. ábra. A tengeri liliomok (Crinoidea) felépítését egy németországi jura példányon mutatjuk be
A gerincesek törzsébe tartozó élőlények általános jellemzője a porcos és csontos belső váz, a gerinchúr illetve gerincoszlop, a hátoldali központi idegrendszer és a hasoldali enésztőrendszer, valamint a két részből álló koponya (agy- és arckoponya).
A gerincesek két nagy csoportra oszthatók: a tágabb értelemben vett halakra (Pisces) és a négylábúakra (Tetrapoda). Utóbbiakon belül különíthető el a kétéltűek (Amphibia), a hüllők (Reptilia), a madarak (Aves) és az emlősök (Mammalia) osztálya.
Kétéltűek osztálya (Amphibia)
A karbonban és permben virágkort élő ősi formák a mezozoikum végén kihaltak. Legismertebb képviselőjük az Ichthyostega, amely a csontoshalak és a négylábúk közötti evolúciós láncszem (142. ábra).
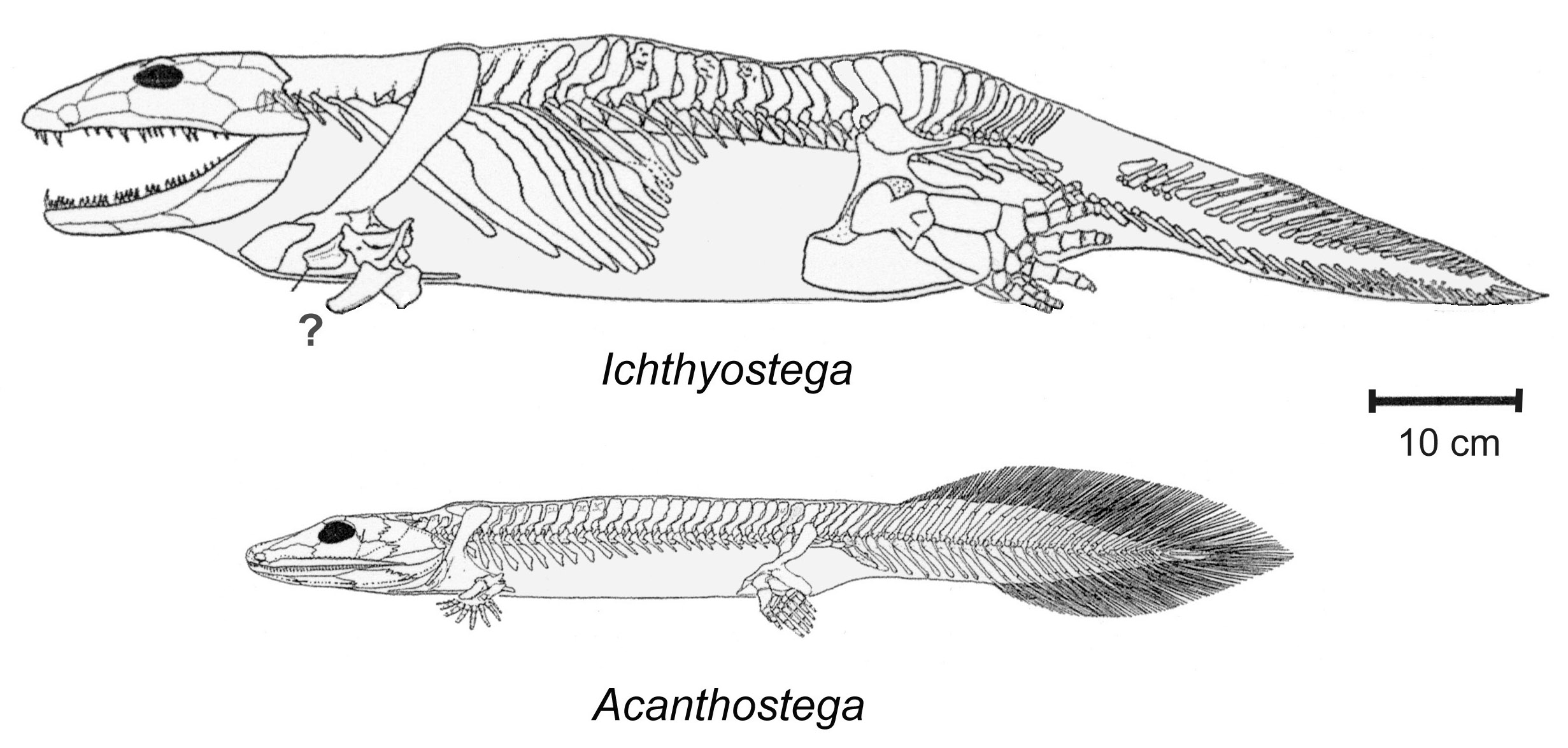 |
142. ábra. A késő-devonban élt ősi kétéltűek (Ichthyostega és Acanthostega) a grönlandi felső-devon rétegekből
Hüllők osztálya (Reptilia)
A gerincesek első igazi szárazföldi osztályát a hüllők alkotják. Nincs lárvaállapotuk és metamorfózisuk, a tojásból kikelt hüllő kicsiny mása a kifejlett példánynak. A tojás kialakulása tette lehetővé a viztől való függetlenedést.
A hüllők rendszerezésében a halántékablakok száma és elhelyezkedése faz egyik legfontosabb bélyeg.
A hüllők földtani-paleontológiai jelentősége elsősorban az evolúció kutatásában mutatkozik meg. Emellett környezetjelzők és utalnak a lemeztektonikai mozgások időbeliségére (faunaprovinciák).
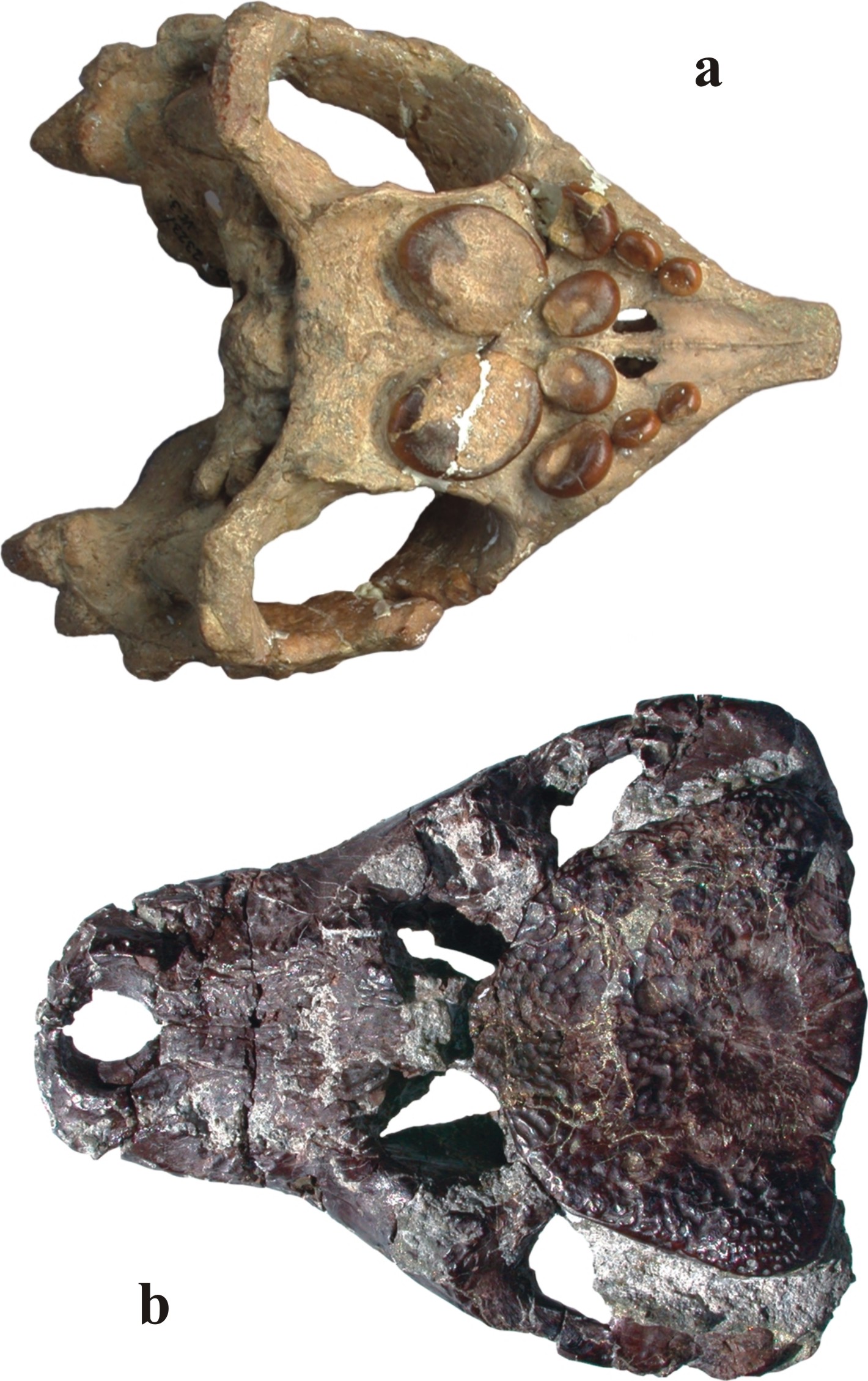 |
143. ábra. A tudományos világban legismertebb magyar fosszília a bakonyi felső-triász rétegekből előkerült kavicsfogú álteknős
Madarak osztálya (Aves)
A madarak a két halántékablakkal ellátott koponyájú, madármedencéjű (ornitischia) hüllőktől származnak. Ritka leleteik szerint a jura időszakban fejlődtek ki (144. ábra). A kréta rendszerből származó példányokon már a mai madarak minden bélyege felismerhető.
 |
144. ábra. Az egyik leghíresebb ősmaradvány a solnhofeni jura Archaeopteryx. Galamb méretű volt és mind a hüllőkre, mind pedig a madarakra jellemző bélyegeket is hordozott
http://www.youtube.com/watch?v=ocr9UaetlbY
Emlősök osztálya (Mammalia)
Az emlősök fő jellemzője a nagy agykoponya, az összetett őrlőfogak, egy csontból álló alsó állkapocs, korlátozott növekedés az egyedfejlődés során, melegvérűség (endotermia), szőrtakaró, elevenszülés, ivadékgondozás. Egyes nézetek szerint nem külön osztály, hanem a hüllőkön belül magas fejlettségi szintet elért, különböző származási vonalon hasonlő tulajdonságokra szert tett (polifiletikus) csoport.
A szerves anyag Földi körforgásában a növények fontos láncszemet alkotnak. A klorofiltartalmú autotróf növények a szervetlenből szerves anyagot állítanak elő, amellyel a növényevő, majd a ragadozó állatok táplálkoznak. A szerves anyagot azután a heterotróf baktériumok (és alárendeltebb mértékben a gombák is) az egyszerű ásványi molekulák szintjére bontják le (foszfát, nitrát stb). A növények szervesanyag-termelése fokozódott, és ezt az állatvilág egyre több lépcsőben, egyre áttételesebben hasznosítja.
A növények és az állatok törzsfejlődésének szoros kapcsolatát különösen jól tükrözi a virágos növények és a rovarok törzsfejlődése (koevolúció). A szerves élet kialakulásakor az ősóceán és az őslégkör oxigéntartalmát kizárólag a sekélytengeri növények, nagyrészt a lebegő életmódot folytató moszatok életműködése szabta meg. Azokban a törzsfejlődési szakaszokban, amikor a fitoplankton feldúsult, az óceánok pH-értéke emelkedett, a CO2-nyomás csökkent, több mésziszap rakódott le, és megnyílt a lehetősége a kőolaj felhalmozódásának. A fitoplankton működésének csökkenésekor viszont a CO2-tartalom növekedett, a pH értéke alacsonyabb lett, és a karbonátos üledékek helyébe kovaüledékek léptek. Ilyenkor a zátonyalkotók és a ragadozók száma csökkent. Az óceánokban a fitoplankton szabályozó szerepe ma is jelentős. A szárazföldi növények szilurbeli elterjedés után a légkör oxigéntartalmának szabályozásában növekvő szerepet játszanak.
3,5 milliárd éve jelentek meg (Warrawoona Csoport, Ausztrália). Az általuk képzett bioszediment struktúrák a sztromatolitok, amelyek fosszilizálódnak. A kékmoszatok a prekambrium jellegzetes maradványai. Földtörténeti-evolúciós jelentőségük az oxigéntermelés miatt rendkívül nagy volt. Extrém, pl. túlsós körülmények között is megélnek.
A sztromatolitok a magyarországi sekélytengeri karbonátos üledékekben meglehetősen gyakoriak. A Dunántúli-középhegység triász platformkarbonátokból felépülő, ciklusos, sekélytengeri rétegsorában az árapályövben képződött sztromatolitok (lofer-ciklus B-tag) összvastagsága több száz méterre tehető (145. ábra).
 |
A moszatok valódi sejtmaggal rendelkező vízinövények, amelyek közül a szilárd vázat építőknek van földtani jelentőségük.
 |
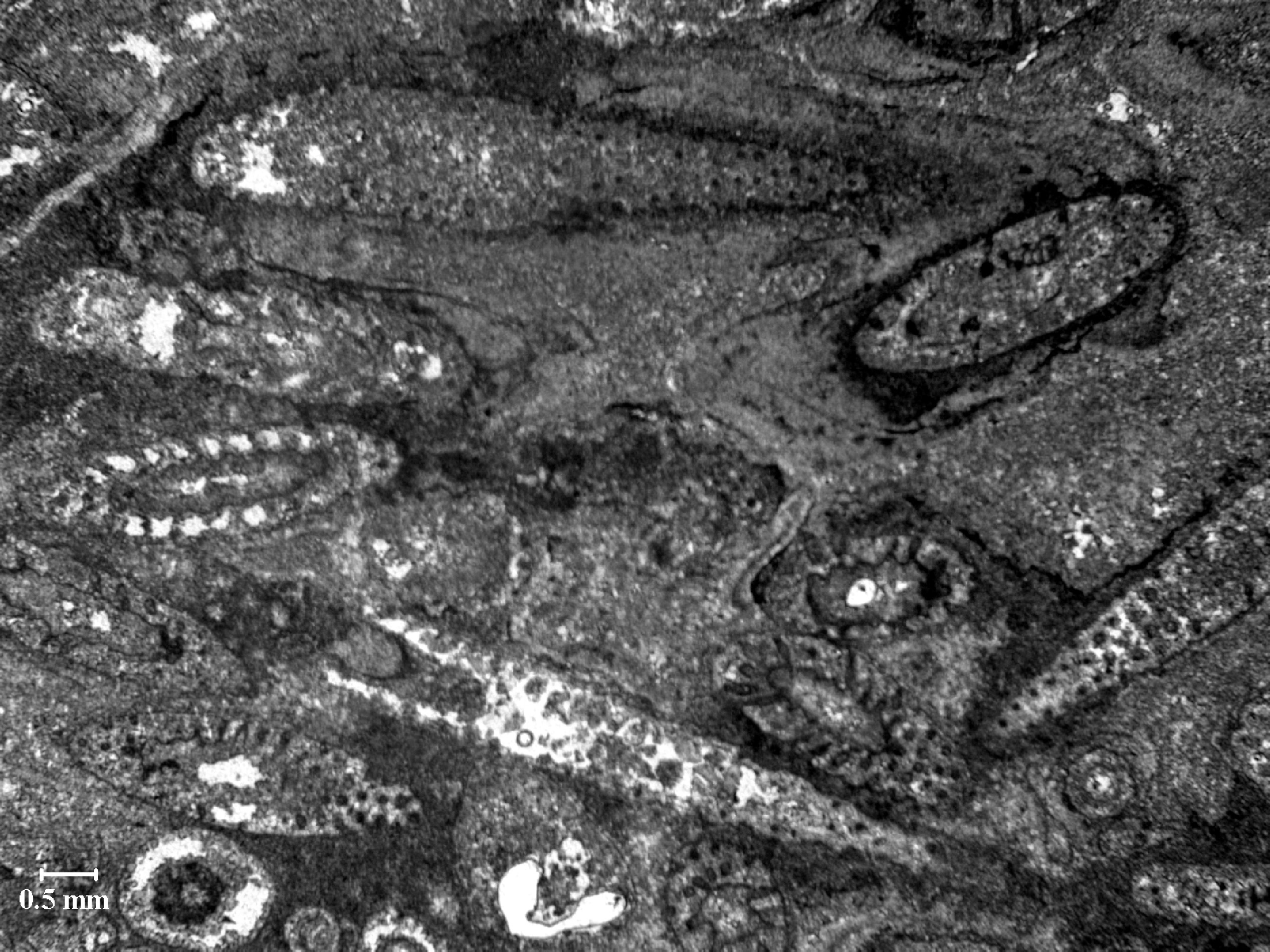
146. ábra. Kokkolitok elektronmikroszkópos felvétele
A sárgásmoszatok közül egyedül a mészpikkelyesek jelentősek földtani szempontból. Ezek vázelemei a változatos alakú kokkolitok, amelyek kőzetalkotó mennyiségben dúsulnak fel egyes tengeri üledékekben (pl. a kréta korú írókréta anyagának 70%-a kokkolitokból áll). A kokkolitok (146. ábra.) egyes csoportjai korhatározásra alkalmasak, segítségükkel a kainozoikumot nannoplankton zónákra lehet tagolni.
A zöldmoszatok (147. ábra) elterjedt, változatos környezetekhez alkalmazkodott csoportjai közül földtani jelentőségük azoknak van, amelyek mészvázat választanak el.
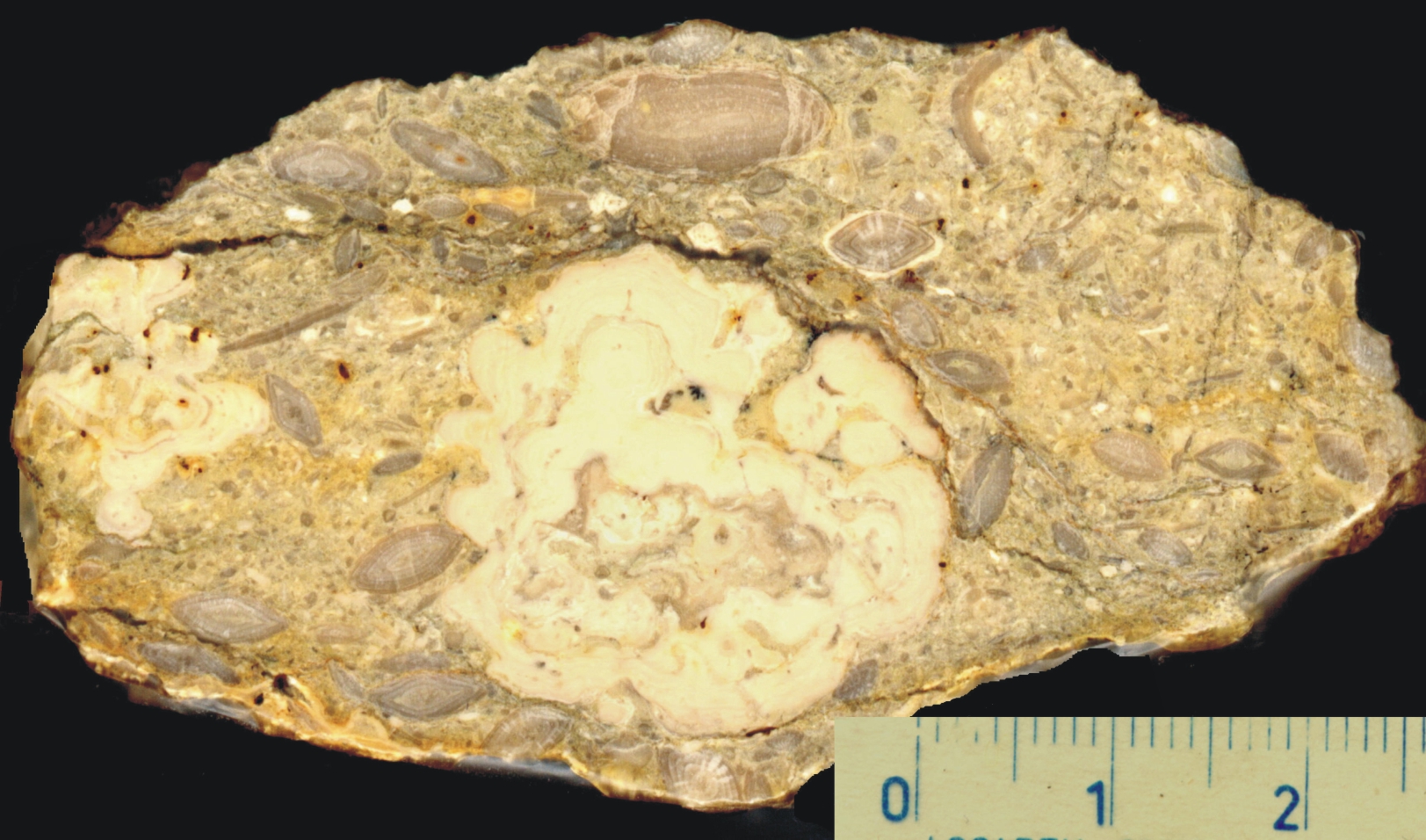
 |
147. ábra. Zöldalgák (Diplopora) vékonycsiszolati képe a bakonyi Tagyoni Mészkőben
 |
148. ábra. Vörösalga-gumó középső-eocén vértesi nummuliteszes mészkőben
A vörösmoszatok túlnyomó részben sekélytengeri környezetben élő, az aljzaton rögzült élőlények. Jelentős szerepet játszanak a sekélytengeri zátonyok külső falának a felépítésében (148. és 149. ábra).
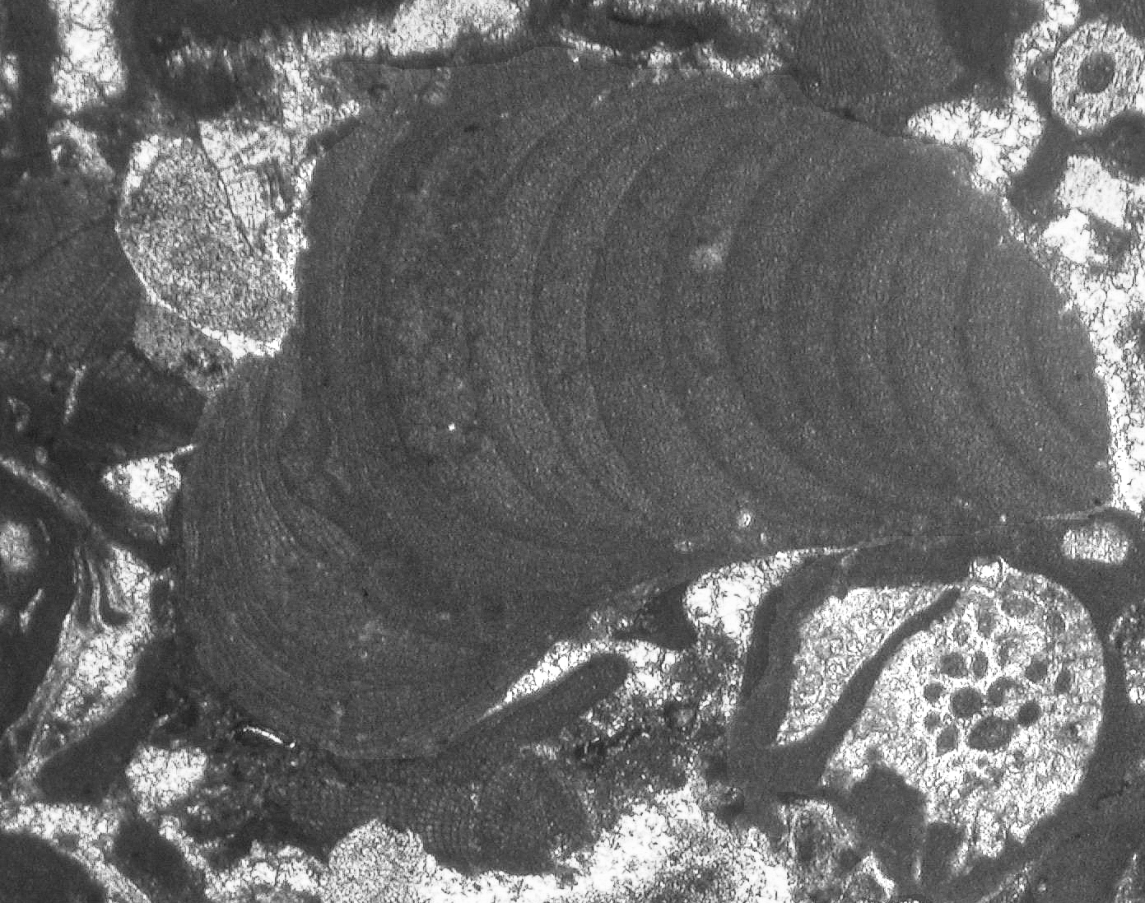 |
149. ábra. Vörösalga vékonycsiszolati képe.
A harasztok az első, szállítószövettel (150.) rendelkező magasabb rendű szárazföldi növények, amelyek szaporodása azonban még vízhez kötött. Megjelenésük a késő-szilurban, adaptív radiációjuk a devonban volt. Ide tartoznak az ősharasztok, a korpafüvek, a zsurlók és a páfrányok.
 |
150. ábra. Lepidodendron (bal oladal) és Sigillaria (jobb oldal) jellegzetes levélripacsai
Az örvös felépítésű zsurlók a devonban jelentek meg, virágkoruk a karbonban volt, de még a triász időszakban is viszonylag gyakoriak voltak. Legismertebb képviselőjük a zsúrlófa (Calamites), amelynek maradványai a Mecsek környéki karbon üledékekből is előkerültek.
Az első magvas növények a nyitvatermők voltak. A devonban jelentek meg, virágkoruk a mezozoikum volt. Lágyszárú formák nem alakultak ki közöttük. Nyolc osztályuk közül kettő földtanilag jelentéktelen. A kihalt magvaspáfrányok jól ismert képviselője a Glossopteris, amely valamennyi déli földrészen elterjedt jellegzetes, hidegtűrő növénytársulást alkotott a késő-paleozoikumban. A Gondwana nevű déli őskontinesnek létezésének éppen a Glossopteris-flóra elterjedése volt az egyik első bizonyítéka.
A cikászok és bennettiteszek a karbonban jelentek meg, virágkoruk a mezozoikumban volt (a benettiteszek a kréta végén kihaltak). A toboztermő tűlevelűek közé tartoznak a ciprusok, ezek képviselőit találták meg Bükkábrányban (7.).
 |
151. ábra. Megkövesedett késő-miocén mocsárciprus-erdő a bükkábrányi lignitfejtésben
Zárvatermők törzse
A zárvatermők a legfejlettebb növények. Zárt magházból kialakult termésük van. Közülük a lágyszárúak a fejlettebbek, csaknem az egész szárazföldet meghódították (pl. a fűfélék). Első biztos előfordulásuk a kora-krétából ismert.
A növényvilág legutóbbi nagy átalakulása a mezozoikum végén történt, azóta új osztály nem jelent meg a Földön.
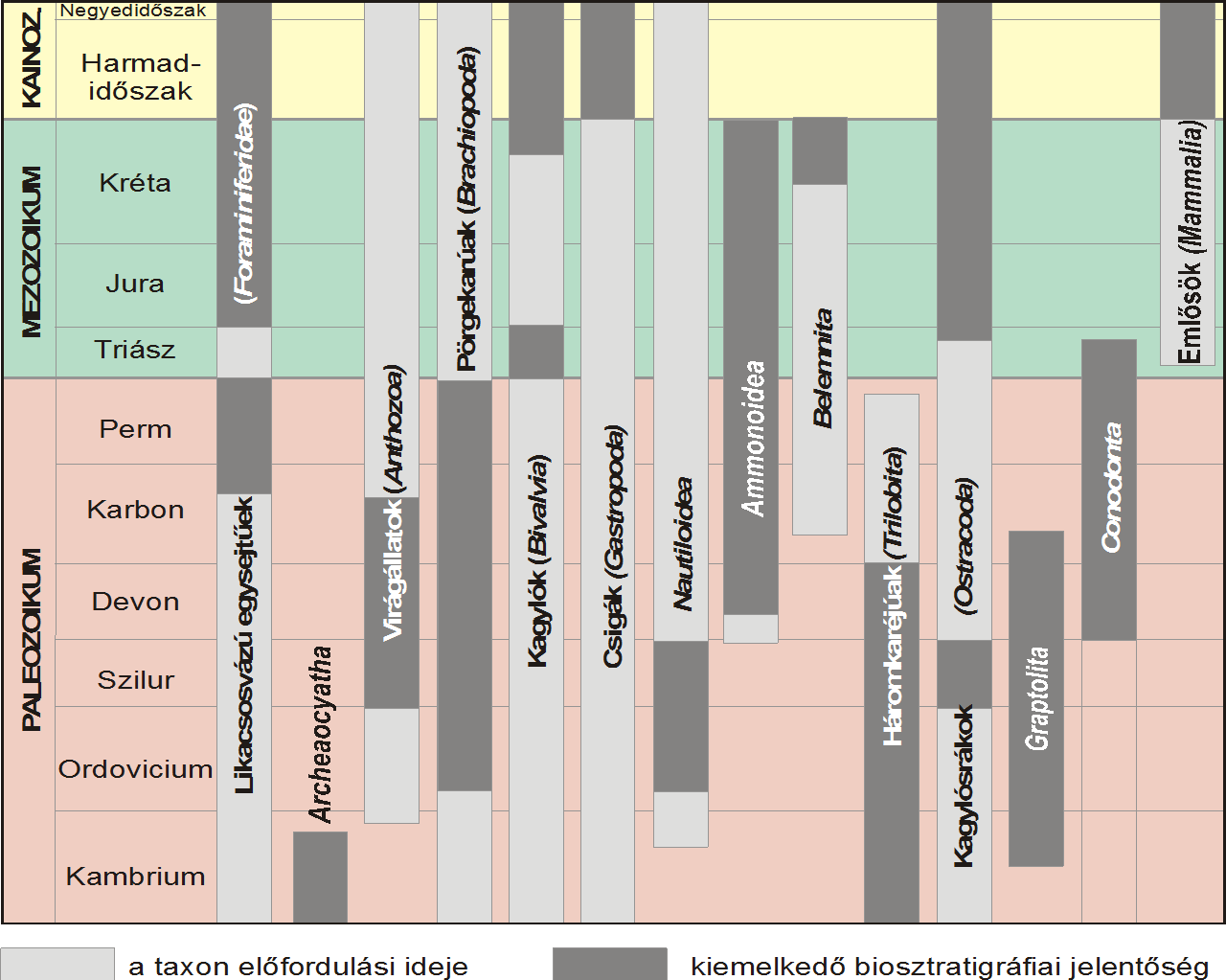 |
152. ábra A legfontosabb korjelző fosszilia-csoportok időbeli előfordulása.