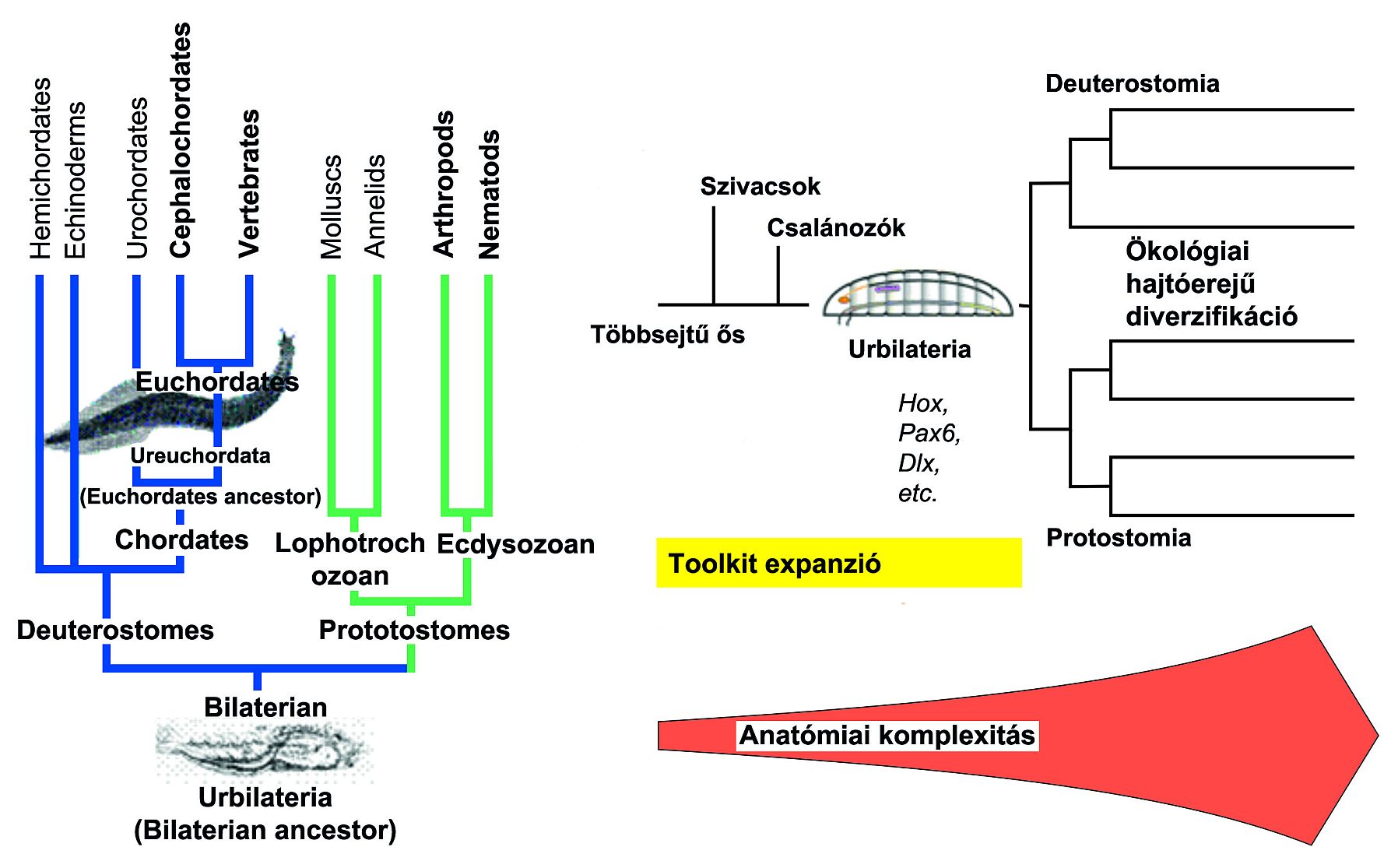
Egyáltalán nem valószínű, hogy előkerül közös ősünk az ősszájúakkal. Ez az elméleti élőlény az Urbilateria vagy PDA (protostome-deuterostome ancestor) minden bizonnyal kistermetű lehetett, megkövülésre alkalmas részeket biztosan nem tartalmazott - miután sem külső, sem belső váza nem valószínű, hogy lett volna, biztosan nem fosszilizálódott jól! 28. ábra
Egy lehetőség mégis maradt arra, hogy komoly következtetéseket vonjunk le rá vonatkozólag: meg lehet vizsgálni azokat a géneket, amelyeknek ugyanaz a funkciója ősszájúakban és ősszájúakban, hiszen aligha tévedünk, ha feltételezzük, hogy ez csak úgy lehetséges, hogy közös ősünkben is ugyanaz lehetett ezeknek a géneknek a funkciója. A kortárs tudomány egyik kiválósága, Sean Carroll nyomán mondhatjuk, hogy ez „paleontológia kövületek nélkül (”paleontology without fossils”).
A közös Bilateria ős lehetséges tulajdonságait elsődlegesen az ízeltlábúakban (Drosophila) és a gerincesekben (egér) egyaránt megtalálható konzerválódott génekből és azok fejlődéssel kapcsolatos funkcióikból következtették ki.
Általánosan elfogadott nézet, hogy a rovarok ventrálisan elhelyezkedő és a gerincesek dorzálisan elhelyezkedő központi idegrendszere párhuzamos fejlődés eredménye. A "dorzális" és "vertralis" kifejezések azonban nem bírnak jól körülírt morfológiai értelemmel, így mindenképpen indokolt az a nézet, hogy referenciarendszerként a korai embrióknak a fő testtengelyek kialakításában résztvevő génjeinek kifejeződési mintázatait használják. A Drosophilánál a dorzálizáló funkciókat a dorzálisan kifejeződő decapentaplegic (dpp) gén kódolja, míg a neki megfelelő (=homológ vagy orthológ) gerinces gén, a BMP-4 ventrálisan fejeződik ki, és ventrálizáló funkciókkal bír. Hasonlóképpen: a Drosophila shor gastrulation egy ventrálizáló faktort kódol, míg Xenopus chordin génnek dorzálizáló hatása van. Az ortholog géneknek ez a dorzális/ventrális kifejeződési rendje vezetett német kutatókat arra a következtetésre, hogy ami "ventrális" a rovaroknál, megfelel a "dorzális"-nak a gerinceseknél, és igy központi idegrendszereik homológ strukturák.
A gerincesek és rovarok központi idegrendszerének kialakulása és a korai testterv gének
A rovar embriók ectodermájának neurogenikus régiója későbbi ventrális, míg a gerinceseké a dorzális oldalon van. Ha azonban a dorsoventrális mintázat kialakításában résztvevő orthológ gének kifejeződését vesszük figyelembe, a neurogenikus régiók mindkét csoportban azonos oldalon helyezkednek el. A neurogenikus régióban mindkét csoportnál azonos csoportba (az achete-scute génkomplex génjeihez hasonló) un. proneurális gének segítik elő az idegsejtek fejlődését, az is nagyon hasonló mechanizmussal dől el, mely sejtekből lesznek neuroblastok a régión belül. A sejtek régión belül elfoglalt poyicióját szintén orthológ gének azonositják az ectodermán belül.
Mind a rovaroknál, mind a gerinceseknél a központi idegrendszer természetes módon felosztható anteriorális és posteriorális részre: Drosophilában azokat a neurogenikus régiókat, amelyekből az anteriorális syncerebrum (a proto-, dento-, és tritocerebrum) alakul ki, rés választja el a posteriorális gnathocerebrumtól. A gerincesekben befűződés választja el az anterioralis pros- és mesencephalont és a posterioralis rhombencephalont. Mind a rovaroknál, mind a gerinceseknél az anteriorális részek az orthodenticle ortholog gének kifejeződési területén alakulnak ki, és az ortholog génekre mutáns egyedekben hiányoznak. A posteriorális részek a Hox-gének kifejeződési doménjén belül alakulnak ki, és a Hox mutánsokban transzformálódnak.
A gerincesek és rovarok agya tehát egymással összevethető anteroposteriorális régiókból alakul ki, ha viszonyítunk az ortholog gének kifejeződési mintázatához.
A hasonló jellegű és funkciójú agyi régiók azonos ortholog géneket fejeznek ki
A vizsgálatok amellett szólnak, hogy filogenetikusan konzervált gének fejeződnek ki az összevethető agyi régiókat. Röviddel a gastrulatiót követően a gerincesek elülső velőpajzsa (neural plate) és a rovarok elülső neurogén régiójának vizsgálata szépen mutatja az elvet. Drosophilában az anteriorális syncerebrum neuroblastjai kifejezik a tailless gént, és a kifejeződésnek megfelelő agyrészek defektívek a tailless mutánsokban. Gerincesekben a tailless-ortholog gén kifejeződése az előagyra korlátozódik. További fontos adalék, hogy mind a Drosophilában, mind az egérben a később kialakuló szemek és az optikai információ átvitelével kapcsolatos agyrészek a tailless-ortholog expressziós doménjébe esnek. Figyelembe véve, hogy a gerinces Pax-6-gén, és annak Drosophila orthologja az eyeless is alapvető szerepet játszik a korai szemfejlődésben, arra gondolhatunk, hogy a gerincesek és rovarok közös őse már fényérzékeny szerveiktől hasonló módon kapta a vizuális bejövő információkat, mint kései utódai. Hasonló horderejű felfedezés, hogy a rovarok és gerincesek agyának egymással könnyen megfeleltethető részei dolgozzák fel a szaglási információt, hasonló stratégiát követve. A szaglással kapcsolatos területeken a Drosophilában, egérben, sőt a Chaenorhabditis elegansban is.
További párhuzamokra is fény derült a vizsgálatok során: A rovarok agyának anteriorális részét, a syncerebrumot különféle neuroblast csoportok alkotják. A leendő tritocerebrumot felépítő neuroblastok korán elkülönülnek a proto- és deuterocerebrumot alkotóktól. A gerincesek agyának anteriorális részén hasonlóan korán különül el az elő- és középagy (prosencephalon és mesencephalon). Mind a rovarok anterioralis szncerebruma, mind a gerincesek prosencephalonja olyan agyrészeket foglal magába, amelyek a memóriával kapcsolatosak (a memóriával kapcsolatos molekuláris szintű folyamatok is nagyon hasonlóak.
A rovaragy posteriorális részében (a gnathocerebrumban) a páros szegmentált ganglionok a gnathális szegmentekből származnak, majd fuzionálnak és kialakítják a subvesophagalis ganglionokat. A ganglionok, együttesen az elülső tritocerebrummal, un. középvonali sejteket vesznek körül. A gerinces nyúltagykezdemény, - akárcsak a gnathocerebrum - szintén alegységekre osztható, az un. rhombdomerekre, amelyek szintén középvonali sejtek számos ortholog gént fejeznek ki. Mint arról már szóltunk, mind a rovarok, mind a gerincesek posteriorális agyrészeinek metamerikus alegységei a Hox-gének specifikus kombinációit fejezik ki.
Hasonló axonelrendeződések a fejlődő rovar- és emlősagyban
Az elsődleges vagy "pionír" neuronoknak egy jól szervezett, egyszerű mintázata alakul a rovarokban és a gerincesekben egyaránt. A neuronok egy korai axonális vázat (=scaffold) alakítanak ki, miközben növekedési kónusaik (=growth cones) az osztodó neuroblastokat körülvevő gliasejtek segítségével tájékozódnak. Az elsődleges axonhálózat még azelőtt kialakul, hogy a magasabb agyi strukturák kialakulnak.
A rovarok gnathocephalonja és ventrális idegkötege axonokból egy létraszerű vázat fejleszt ki, amelyek a szegmentenkénti commissuralis axonkötegeket és két hosszanti köteget alkotnak, amelyek a középvonali sejtektől laterálisan futnak. A gerincesek szintén nagyon kifejezett longitudinalis kötegekkel bírnak, amelyek középvonali sejteket vesznek körül, amelyekből később alaplemez (floor plate), és a commissuralis axonok többnyire a rhombomer határokon kereszteződnek. A metamerikus alegységek páros idegeket fejlesztenek mind a rovaroknál, mind a gerinceseknél.
Az axonkapcsolatok nagyon hasonlóak a syncerebrumban és a gerinces elő- és középagyban is. Rovaroknál az elülső agyrészben középtájon az axonpályák olyan korszerű képletet formálnak, amely körülveszi az előbél kezdeti szakaszát, ill. a leendő infundibulumot és hipotalamuszt a gerincesekben. Az elülső agyi régió rovarokban és gerincesekben is felosztható egy központi részre, amelyet egy axongyűrű vesz körül, és egy perifériás részre, amely a gyűrűn kívül helyezkedik el. Az axongyűrűt mindkét csoportnál kifejezett pályák kötik össze a szemekkel. Mindkét egységre jellemző, hogy a központi részben további gének fejeződnek ki.
A Drosophilában a decapentaplegic gén az előbél anteriorális részén fejeződik ki, míg a vele ortholog BMP-4 kifejeződése a leendő infundibulumra korlátozódik. A stomodeumból fejlődnek ki a stomatogastricus ganglionok, míg a gerinces hipotalamusz az autonom idegrendszer koordinátoraként működik, ideértve a gastrointestinalis mozgások szervezését. Valószínűnek látszik tehát, hogy a gerincesek ventrális diencephalikus régiója, amely magába foglalja a hipotalamuszt és az infundibulumot, megfelel a rovarok stomodealis régiójának, és funkciójuk hasonlósága nem puszta véletlen.
Az elmondottak alapján tehát felvázolható a rovarok és gerincesek közös ősének (a régebbi kifejezést használva „gastroneuralis közös ősnek”) néhány tulajdonsága:
Központi idegrendszere felosztható volt anterioralis és posterioralis részre, amelyekből a rovarok és gerincesek agyának jól elkülöníthető anterioralis és posterioralis részei fejlődtek.
A mai ortholog gének őseinek alapvető expressziós mintázata fennmaradt, így a gerincesek és rovarok ortholog génjei jól megfeleltethető agyrégiókban fejeződnek ki, amelyeknek funkciója is hasonló. A közös ősnek valószínűleg megvoltak a hasonló agyrégiói hasonló funkciókkal. A Hox gének kifejeződési mintázata alapján arra lehet következtetni, hogy metamerikus osztottságú élőlény lehetett.
A középvonali sejtek közös volta alapján arra gondolnak, hogy a közös ősnek egy hosszű blastopórusa lehetett, anterioralis a száj, ill. posterioralis végén az anus. Az idegrendszer talán anterioralisan és a lateralis blastoporus szegély mentén képződött. A központi idegrendszer középvonali régiója talán a laterális blastoporus ajkak összezáródását jelzi. Stomodaeuma ventrálisan helyezkedett volna el. A gerincesek evolúciója során egy D/V tengelyirányváltás következhetett be, és az agy közepén található száj talán eltűnt és egy új száj alakult ki az új ventrális (addig dorzalis) oldalon. A gerincesekben máig fellelhető axon gyűrű talán azt a helyet jelzi, ahol az ősi száj elhelyezkedhetett. A hosszanti axonkötegek talán azt a helyet jelzik, ahol a blastoporus egykor elhelyezkedett.
Végezetül szólunk néhány olyan eredményről, amely közvetve a hipotézis mellett szól. A laposférgek axonális mintázatát tanulmányozva arra a következtetésre jutottak, hogy valószínűtlen, hogy kétszer függetlenül ugyanaz az axonmintázat alakuljon ki. Egy csoporton belül is igen eltérő mintázatok alakultak ki, és nem hasonlítottak sem rovarokéra, sem a gerincesekére.
Az sem valószínű, hogy az egyes gének függetlenül kerültek kapcsolatba ennyire hasonló folyamatokkal. Sokkal valószínűbb, hogy mint szabályozó rendszer együttesen jelen voltak a közös ősben. Ez azt jelenti, hogy a rovarok és gerincesek agya közös alapterv szerint épül fel.
További közös vonások
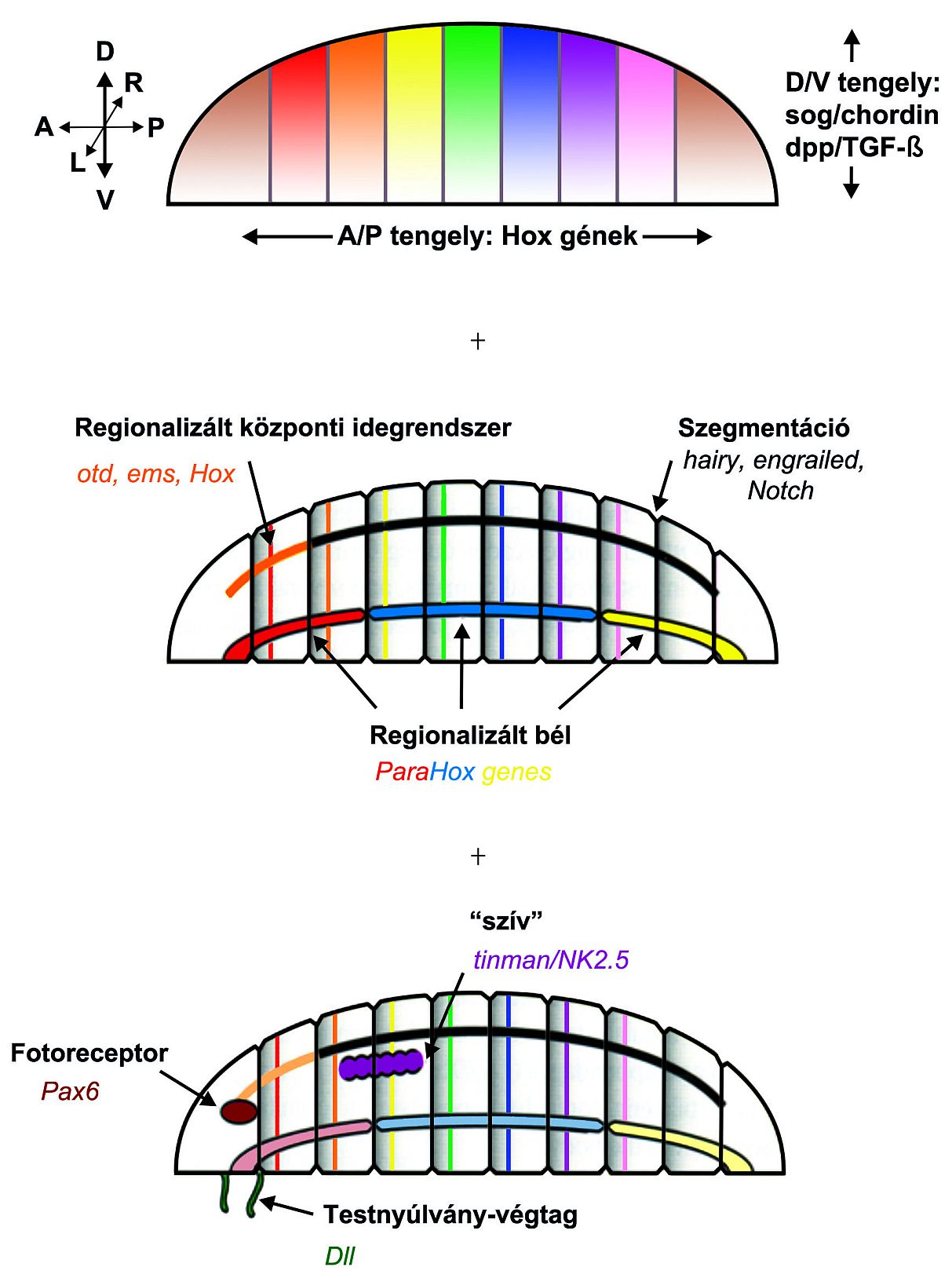
A dorzoventrális tengely kialakítását talán az ősi, sog/chordin és TGF-β géncsaládok irányíthatták. Az anterio-poszterior tengely további felosztását valószínűleg Hox gének kifejeződésének átfedő doménjei végezték. A különböző szövetrétegek régiók szerint helyezkedtek el végig az A/P tengely mentén, beleértve az emésztő/bélrendszert (Parahox gének) és az idegrendszert (otd, ems, Hox).
A szegmentáció talán az ősi hairy, engrailed, Notch gének szabályozása alatt alakult ki. Egy fotoreceptor szerv, egy „szív”, a légzőrendszer (lásd előbb) és egy testfalkitüremkedés "primitív" (a jelző erősen megkérdőjelezhető a csirke és Drosophila végtagjában működő jelátviteli rendszerek azonossága alapján, lásd később) változatai lehettek jelen a Bilateria ősben, (ugyanilyen sorrendben) az ősi Pax 6 (sine oculis, dachshound), tinman és DII gének által szabályozva.
Másik fontos génkészlet, amely közös a protostomatáknál és deuterostomatáknál, azoknak a transzkripciós faktoroknak a génjei, amelyeknek a fej kialakításában van szerepük. Drosophilában az agyat három szegment, neuromer alkotja, amelyeket három transzkripciós faktor specifikál. Génjeik: tailless(tll) és orthodenticle (otd) – a legelső neuromerben, főleg
empty spiracles (ems) - két hátsó neuromerben. Loss of function mutációk a hozzárendelt génekben eliminálják az említett neuromereket. Bicoid és hunchback gradiensek indukálják őket.
Béka és egér homológok: Otx1, Otx2 (az otd-é), Emx1, Emx2 (ems-é)– elülső dorzális mezoderma és entoderma indukálja őket, ami nem csoda, hiszen a bicoid a rovar evolúcióban is viszonylag új jövevény. Az Otx2 „knock out-ja” esetén a rhombomer 3 előtt mezodermális és neuronális deffektusok jelentkeznek.
Az ember - EMX2 mutációi schizencephalia nevű kórképhez vezetnek, ami a teljes cortexet átszelő hasadékokban nyilvánul meg 29. ábra.
Anatómiai hasonlóságok
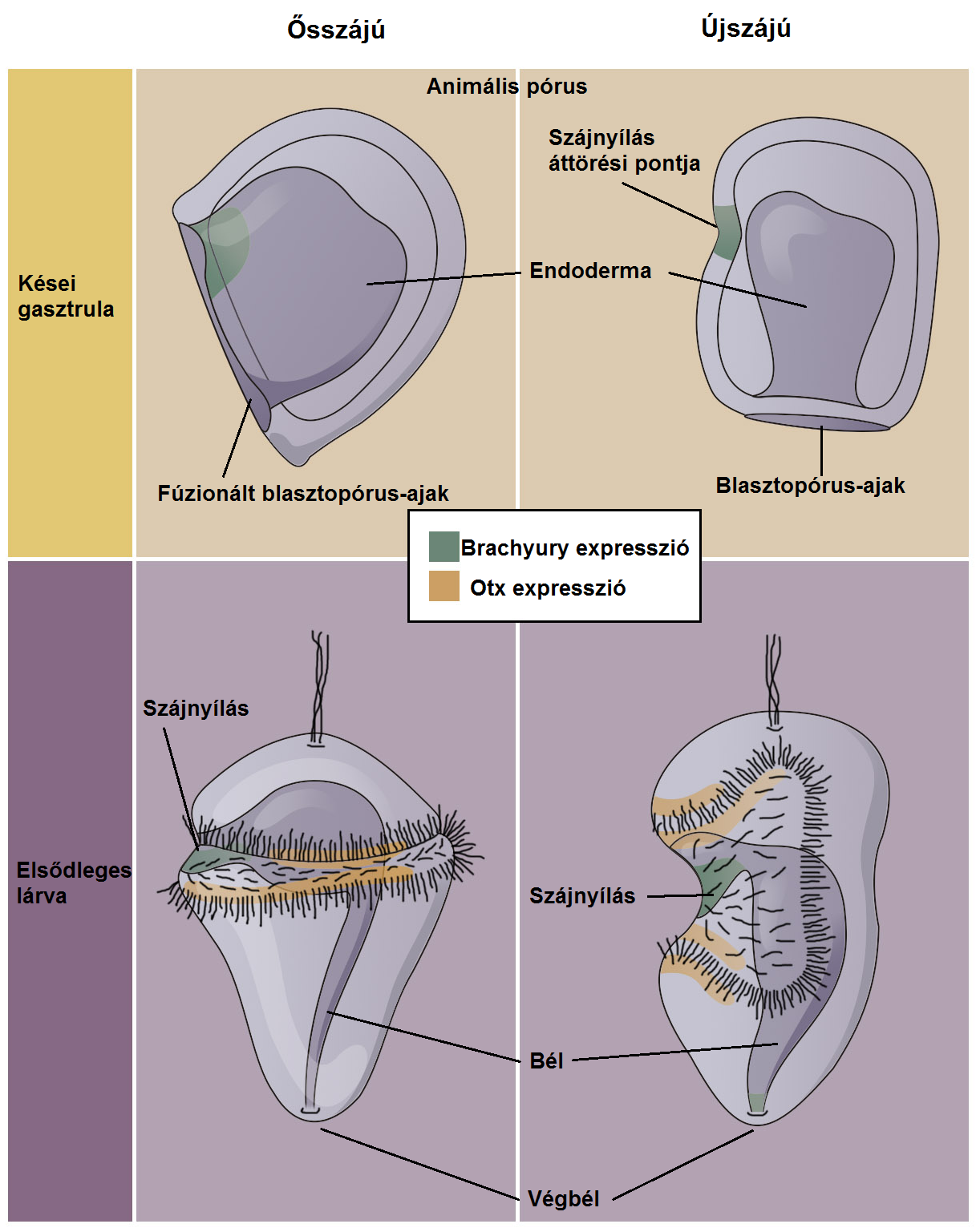
A „legbazálisabb” taxonok csillós lárvákból fejlődnek. A protostomatáknál a blasztopórusból előbél és száj alakul ki, míg a deuterostomatáknál végbél (anus). Újabb eredmény, hogy a Brachyury (T), Goosecoid és Otx gén kifejeződik a pluteus és hemichordata lárvákban és az annelidák trochophora lárvájában is a hasi előbélben – a PDA-tól örökölt csillós lárvákról lévén szó, az eredmény nem is annyira meglepő!
Rendszerint az Urbilateriának egy blasztopórus ajka volt, ami kiterjedt a lárva felszínére, és az ősszajúaknál szájjá, az újszájúaknál végbéllé transzformálódott 30. ábra.
A Cnidariáknak planula lárvájuk van. Bizonyos Cnidariáknál megvannak a harmadik csíralemez (a bilaterális szimmetria előfeltételének) kezdetei - itt van is kezdetleges bilaterális szimmetria! Megvannak: transzkripciós faktorok (snail, Otx, Hox), jelátviteli utak (TGF-β, Wnt), Brachyury a blasztopórus körül fejeződik ki. A Hydra tengelyeinek meghatározásában, akárcsak a komplexebb állatokéban, részt vesz a Wnt-, chordin-, BMP jelátvitel, az Anthozoák Bmp4/dpp homológjai csak a blasztopórus egyik oldalán fejeződik ki, mint bilateráliáknál.
Érdekes felvetés látott napvilágot: Ha Cnidaria a PDA, akkor a testükből lett a bilateráliák agya?
A kérdés nem légből kapott! Meinhardt leírta, hogy míg az a gerinceseknél és rovaroknál Otx- előagyban fejeződik ki, addig a Hydra egész testében, a Hydra posteriorális részén kifejeződő gének pedig a Bilateraliáknál az agy alapjánál.
A bilateriák őse egy radiálisan szimmetrikus, hidraszerű lény lehetett - teste hasonlíthatott a későbbi fejekre - a test később, a háti középvonal mentén történő konvergens extenzióval alakult volna ki az újszájúaknál, új szegmentek hozzáadásával az ősszájúaknál.
A Dll gén kifejeződése a Bilateráiáknál
A DII-es fehérje számos Bilateriánál testfal kitüremkedésekben fejeződik ki, beleértve mind a három fő Bilaterália klád (ízeltlábúak, Lophotrophozoák, és újszájúak) képviselőit.
A DII expresszió jelzi a fej részeit /függelékeit és a lobopoda járólábakat a karmos féreglábú embrióiban, polychaeta/ soksertéjű embrióban a DII fehérje a növekvő parapodiákban és más egyéb függelékekben fejeződik ki, a DII fehérje expresszálódik a csőlábak ill. a tüskék disztális csúcsán lévő sejteken, egy frissen átalakult tengerisünben. A DII expresszió jelzi a proximo-disztális (P/D) kinövések disztális csúcsát ezekben a törzsekben, azt sugallva, hogy a P/D tengely az Urbilateriáknál alakult ki és a DII-es fehérje jelölte ki.
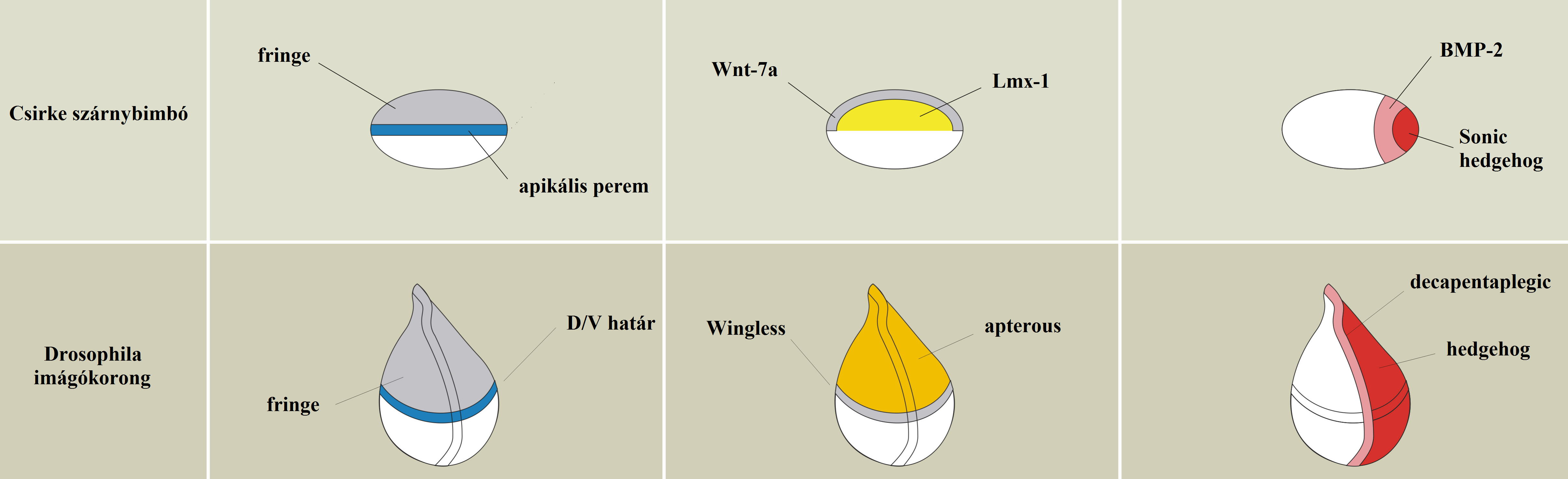
A Bilateriák lábai/végtagjai nem a szó szoros értelmében homológok az Urbilateria függelékeivel, inkább „paralógok”: ősi szabályozási útvonalakat használnak és módosítgatnak újra- és újra. Az egybeesések mindenesetre messze túlmennek azon, hogy a Dll mindenhol előfordul. A jelátviteli útvonalak gyakorlatilag ugyanazok a vizsgált gerinces és Drosophila végtagkezdeményekben, így nyilvánvaló, hogy az Urbilateriának már finoman tagolt végtagfüggelékei lehettek, amelyek kialakulásának hátterében cizellált jelátviteli hálózat állt 31. ábra.
10.31. ábra - Két gyökeresen eltérő végtagtípus - megdöbbentően hasonlító jelátviteli rendszerek szabályozása alatt
 |