A Fajok eredetének megjelenése után sok-sok ideig az elméletet elfogadó kutatók úgy tekintettek az evolúcióra, mint a globális fejlődést, progressziót leíró paradigmára. Ahogy a korábbi felfogások, úgy az evolúciós elmélet is kiemelt helyet biztosít fajunknak: az ember az evolúciós folyamat csodálatos vívmánya, különös tekintettel a nyugat-európaiakra, akik a humán evolúció csúcsai. Darwin maga sem a haladást, sem a „javulást” nem tekintette az evolúció feltétlen velejárójának, de rajta kívül szinte mindenki kritikus tulajdonságnak gondolta a progresszivitást. A következőkben megvizsgáljuk a trendek természetét, lehetséges okát, és hogy van-e értelme evolúciós haladásról beszélni?
A trend definiálható az idők folyamán bizonyos irányba történő változásként. A progresszió, vagy haladás magában foglal bizonyos „javulást”, így feltételez értékítéletet is, azaz hogy eldönthető, mi is a „jobb”.
Trendek: fajták és okok
A trend egy sajátság átlagának, esetleg maximumának vagy minimumának folytatólagos, bizonyos irányba történő változása egy kládban az idők folyamán.A filogenetikai szempontból lokális trend egy egyedi klánra korlátozódik, míg a globális trend az egész élővilágra vonatkozik. A trendeket feloszthatjuk aktív és passzív trendekre.
Passzív trendről beszélünk, ha a kládok vonalai mindkét irányba egyenlő eséllyel fejlődhetnek, de ha az egyik oldalon egy áthághatatlan akadály tornyosul, a vonalak közötti variáció csakis a másik irányba terjedhet ki, ezzel együtt a középérték és a maximum is ugyanerre tolódik. Azzal együtt, hogy a középérték nő, bizonyos vonalak megmaradnak az ősi érték közelében, a minimális méret nem változik. Példa a passzív trendre az emlősök testméretének növekedése. A paleontológusok már régen észrevették, hogy sok állatcsoportban tendencia van a testméret növekedésére, ez a Cope-szabály néven ismert trend. Nagyszámú (1534) észak-amerikai krétakori és újkori emlős testméretének analízise ilyen passzív trendet mutat. Az emlősök kis testméretűek voltak a kréta-tercier (K/T) határon történt tömeges kihalás után, és a minimális testméret számottevően nem változott. Az átlag és a maximális méret növekedett, különösen a K/T-kihalás után, amikor az emlősök diverzitása robbanásszerűen nőni kezdett. Amikor 779 (minden bizonnyal) ős-leszármazott fajpárt vizsgáltak, túlnyomó részt testméret-növekedést tapasztaltak. Ez a statisztikus eltérés természetes szelekcióra utal.
Az „irányított” (driven) trendnél a vonalon belül az egyik irányba történő változás valószínűbb, mint a másikba, a minimum és maximum is változik az átlag változásával. A lovak némileg kilógnak az emlősök közül: nemcsak testméretük maximuma és átlaga, hanem minimuma is növekszik az újkorban, az ős-leszármazott párok is gyakrabban mutattak növekedést, mint méretcsökkenést. Mindez „irányított” trendre utal.
Legalább négy légy csoporton belül volt kimutatható a fej és a szem kiszélesedése 9. ábra.
6.9. ábra - A Zygothrica csoporton belül (Drosophilidae) az evolúció a szélesebb szemek és fejek irányába haladt, amit a filogenetikai analízis is igazol
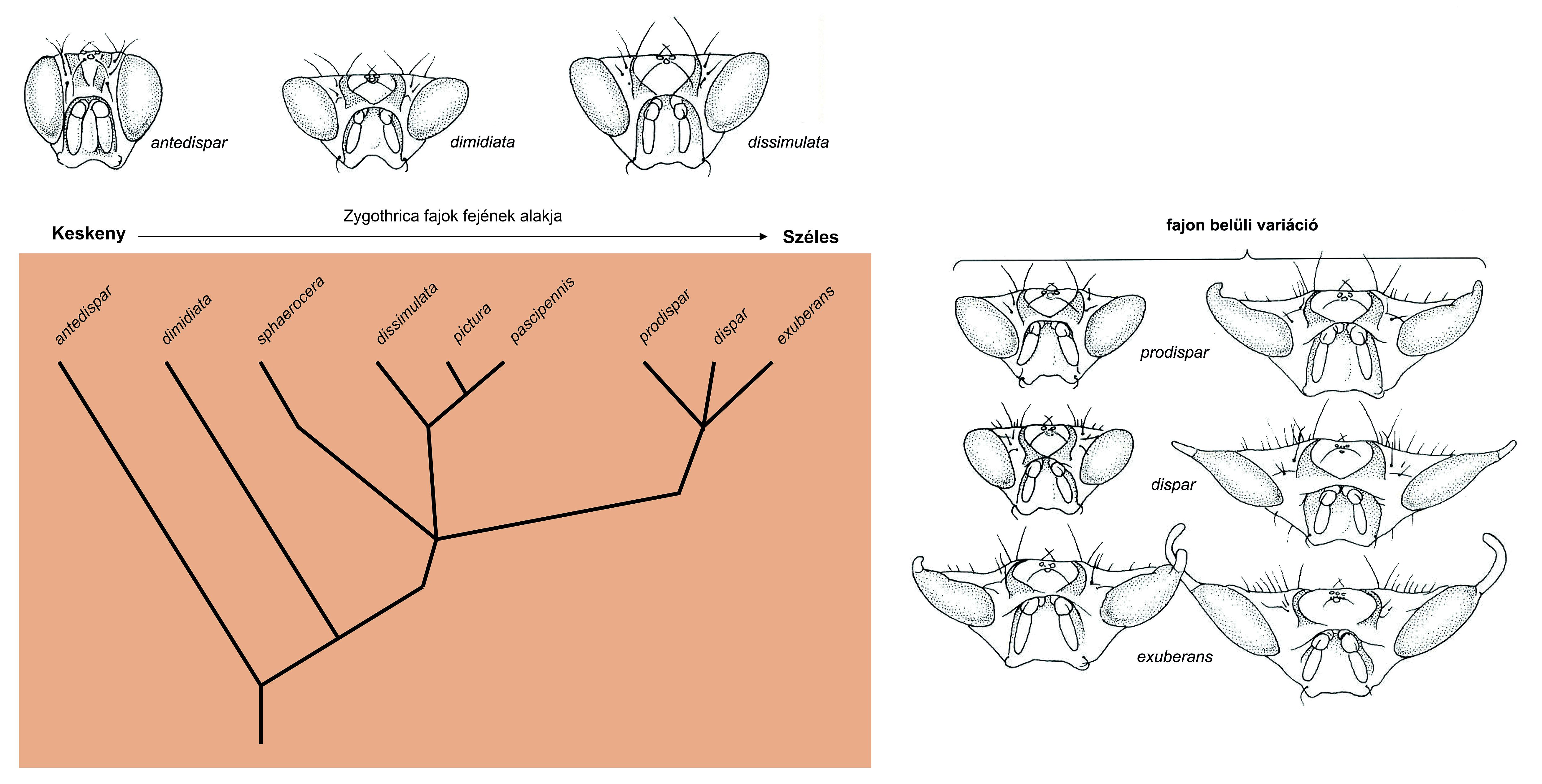 |
A fenti trendek az egyedekre irányuló szelekciónak tudhatók be. A fajszelekciónak köszönhető trend szép példája a nem-planktotrofikus fajok arányának emelkedése a planktotrofikusakhoz (azaz planktonikus lárvaként táplálkozókhoz) képest több újkori csiga-kládban. Azok a fajok, amelyeknek nincsen planktotrofikus diszperziós életszakaszuk hajlamosabbak a kihalásra, amit messzemenően ellensúlyoz magasabb fajképződési rátájuk. A magasabb fajképződési ráta annak köszönhető, hogy alacsonyabb diszperziós rátájuk lecsökkenti a génáramlást a populációk között.
Ha bármilyen jelleg elősegíti, hogy egy klád fajokban gazdagabbá váljon a jellegnek a speciációra vagy a kihalásra gyakorolt hatása által, a klád minden más jellege magasabb frekvenciával jelenhet meg. A felnőtt rovarokra jellemző felcsavarodott szívó szájszerv azért vált kiemelten gyakorivá, mert jellemző vonása a szélsőségesen változatos Lepidopteráknak. A Lepidoptera lárvák növényevők, és a növényevő sajátság következetesen kapcsolódik a magas diverzifikációs rátákhoz a rovaroknál. A szívó szájszerv megemelkedett gyakorisága valószínűleg annak köszönhető tehát, hogy „együtt ment” (az angol nyelvű szakirodalom a „hitchiking” azaz (autó)stoppolás kifejezést használja) a növényevő sajátsággal. Ha egy jelleg speciációs vagy extinkciós ráta különbséget okoz, vagy pusztán az ilyen jelleggel korrelációt mutat, a jelleg „vele megy” a keletkező fajjal (species hitchhiking).
A passzív trendeket kikényszerítő korlátok funkcionális, vagy fejlődésgenetikai természetűek lehetnek. A legkisebb madarak és emlősök például valószínűleg elérték a testméret funkcionális minimumát, mivel kisebb méretű állat valószínűleg nem tudná fenntartani a magas testhőmérsékletet a már túl nagy felület/ térfogat arány miatt.
Kialakulhatnak továbbá olyan fejlődési útvonalak, amelyek reteszekként (angolul: ratchet) szolgálnak, olyan mechanizmusokként, amelyek a visszaalakulást valószínűtlenné teszi. A rovarok torának például minden szelvénye kiegyénült, és nem valószínű, hogy újra azonos morfológiájúvá válhatnak, mint őseikre jellemző lehetett. A legszélsőségesebb epigenetikai reteszek komplex tulajdonságok elveszésénél lépnek fel, amelyek ugyanolyan formában valószínűtlen, hogy még egyszer kialakulhatnának; ez Dollo törvénye. Bizonyos ilyen evolúciós változások, úgy tűnik, irreverzibilisek: sohasem láttak még arra példát, hogy tojásrakó ősökkel rendelkező elevenszülő gyíkok és kígyók utódfajai újra tojásrakókká váljanak.
Vannak olyan trendek vagy irányok, amelyek az élet egész történetére jellemzőek?