Tartalom
- Kapcsolat a mikro- és makroevolúció között
- Evolúciós mintázatok, makroevolúció
- Az állatok leszármazási fája
- Fosszíliák - elsődleges források az élet történetének megértéséhez
- Evolúció és testtervek
- A makroevolúció előfeltételei
- Molekuláris parszimónia
- Heterokrónia - életciklusváltozások
- „Tinkering” az anatómiai struktúrák szintjén: a gerincesek zárt vérkeringési szervrendszerének anatómiai egységei
- „Tinkering” (barkácsolás) a gének szintjén
- Heterotópia
- Heterokrónia II.
- Heterometria
- Heterotípia
- A barkácsolás egy szélsőséges példája: a kromatin és az éjszakai látás
Az evolúció természetesen egy és oszthatatlan. A biológusok azért használják a makro-, és mikroevolúció szakkifejezéseket, hogy könnyebb legyen eligazodni az evolúciós mintázatok és folyamatok hatalmas tárházában. Az evolúciós folyamatok alapvetően két szintre oszthatók: faj alatti és feletti szintekre.
A mikroevolúció a faj populációin belül („faj alatti szinten”) történő változásait foglalja magába: populációk variabilitása, adaptív változások, fajképződés felé vivő mechanizmusok tartoznak elsősorban ebbe a témakörbe. Fontos sajátsága, hogy rövidebb időtávon értelmezhető.
Ezzel szemben (?) a makroevolúció felöleli a faj szintje feletti változásokat, ide értve a speciációt, és a magasabb rendszertani egységek kialakulását, egészen a törzsek szintjéig, ide tartozó témakörök még az evolúciós újdonságok, új adaptív zónák meghódítása, evolúciós ráták, (esetleges) trendek kialakulása, korlátok kialakulása és a fajok közötti interakciók. Hosszabb időtávon értelmezhető. A makroevolúció, amely erőteljesen átfed az evolúciós fejlődésbiológiával (EVO-DEVO-val), így, egyfajta csúcspontjaként tárgyaljuk az evolúció és fejlődés kapcsolatát.
Fontos megjegyezni, hogy nem minden, sokszor mégoly látványos evolúciós változás vezet új fajokhoz.
A kifejezésekben a biológiai evolúció és diverzitás tanulmányozásának két alapvető megközelítése tükröződik. Az élő organizmusokat tanulmányozó biológusokat korlátozza saját- és a tanulmányozott élőlények életideje: még az ideális kísérleti organizmusok, mint a Drosophila melanogaster, sőt a Caenorhabditis elegans, amelyeknek életideje napokban mérhető, is csak korlátozott számú generáción keresztül tanulmányozhatók, ami finom beosztású, mikroevolúciós skálát jelent. A paleontológusok munkájának korlátai a kövületek korából adódik, ami hatalmas időtávlatot, akár milliárd éveket jelent. Ez egy durvább beosztású, makroevolúciós skálát jelent. A makroevolúció fogalmat - minden bizonnyal ugyanilyen értelemben - Jurij Filipcsenkó 1. ábra orosz biológus használta először, 1927-ben.

A skálák közötti gyakorlati különbségtétel nem könnyű, kérdéses, hogy egyáltalán lehetséges-e közmegelégedésre: sok makroevolúcióval foglalkozó szakember felfogása szerint tudományterületüknek saját, sajátos folyamatai vannak, nem pusztán ugyanazok a folyamatok és mintázatok ismétlődnek hosszabb ideig, mint a mikroevolúciónál. Mások csakis a mikroevolúciós folyamatokat fogadják el az evolúciós változások alapjául, szerintük a fajon belüli változatosság minden evolúciós változáshoz elég nyersanyagot szolgáltat.
Steven J. Gould (2. ábra) , a magyar felmenőkkel is rendelkező kiváló evolúcióbiológus más nézőpontot javasolt, szerinte pluralisztikusabb szintézis szükséges. Ez a szintézis nem veti el a makroevolúciót, vagy vonja kétségbe annak jelentőségét. Arra törekszik, hogy –kiterjesztve az addigi látókört – folyamatok egy hierarchiájának, vagy még inkább hálózatának rendszerét ölelje fel: ide tartoznak a populációkon belül, a fajok között ható, és a fajok szintje felett ható folyamatok is.

Georg Gaylord Simpson (1. ábra) számára évtizedekkel ezelőtt világos volt, hogy széles alapokról kell építkezni. Olyan, nagyobb taxonómiai egységek eredete érdekelte, mint a madarak, vagy a virágos növények. Simpson eszméje volt a kvantumevolúció: az a lehetőség, hogy a fajokon belül fellépő, fokozatos változásoktól gyökeresen eltérő, gyors és drasztikus biológiai változások jöhetnek létre. Simpson munkatársa, Ernst Mayr (1. ábra) kételkedett a kvantumevolúcióban, úgy gondolta, hogy a fajképződési folyamatok gyorsan játszódnak le a kis, helyi populációkban, de a változások csak a paleontológusok szélesebb perspektívájából tűnnek azonnalinak. Végeredményben ezek a látszólag ellentmondó eszmék fejlődtek a 70-es évek szaggatott egyensúly eszméjévé, amely Niles Eldredge és Steven J. Gould (2. ábra) nevéhez fűződnek.
Simpson és Mayr kortársai legtöbben megkerülték a problémát: a mikro-, és makroevolúció közötti rést, Darwin nyomán - aki mindemellett teljesen átérezte, milyen fontosak a kövületek az evolúció szempontjából - a fosszilis anyag hiányosságainak tudták be az ugrásszerű változásokat. Ezen nézőpont szerint más mechanizmusokra, mint a Darwin által javasolt, természetes szelekcióval zajló fokozatos adaptív evolúcióra egyáltalán nincsen szükség.
Az ösztöndíjasként a Szovjetunióból az USA-ba ragadt Dobzhansky (1. ábra) volt a legkiválóbb képviselője annak a nézetnek, hogy a mikroevolúciós változások magukban elégségesek az evolúció teljes körű magyarázatára. A modern, vagy neo-darwiniánus szintézis egyik atyja. A makroevolúció névadójaként említett Filipcsenkó mentora volt a fiatal Dobzhanskynak, és sajátos csavar a történetben, hogy bár gyakran említik érdemeként, hogy ő vezette be az angol nyelvterületre a makroevolúció fogalmát, ezt azért tette, hogy könnyebben érveljen ellene, azaz tagadja a mikroevolúció és makroevolúció közötti különbséget. 1937-es nagy művében, amelynek címe: Genetics and the Origin of Species, szavai szerint vonakodva, de egyenlőségjelet tesz a makro- és mikroevolúció mechanizmusai közé.
Azok a kutatók, akik nem értenek egyet a „makroevolúció= mikroevolúció hosszú távon” nézettel (a neo-darwini szintézissel), úgy szoktak érvelni, hogy ez a felfogás nem ad elfogadható magyarázatot az olyan lényegi evolúciós újításokra, mint a rovarok szárnyai, vagy a négylábú gerincesek lábai. A nagyobb egységek eredete és az evolúciós újítások kutatása a huszonegyedik század elejétől reneszánszát éli: a technikai arzenál eddigre érte el azt a szintet, hogy tanulmányozásuk egy minőségileg új megközelítésben történhetett, és a szemléleti változás is sokat segített a klasszikus problémák megoldásában. Egységbe forrtak a klasszikus populációs genetikai, klasszikus szisztematikai, paleontológiai módszerek a molekuláris szisztematikával, molekuláris evolúcióval és a fejlődésgenetikával. Kiterjesztett fegyvertárukkal a makroevolúció kutatói egyre szigorúbb tesztelésnek tudják alávetni invenciózus, de korábban meglehetősen spekulatívnak számító elméleteiket.
A mikroevolúció tárgya az egyedek változatossága a populáción belül, és a variabilitás az idők folyamán történő változása. Az egyedek által hordozott allélok az egymásra következő generációk során gyakoribbakká vagy ritkábbakká válhatnak. Alapvetően két mikroevolúciós folyamatnak köszönhetőek az ilyen változások: a természetes szelekcióval folyó evolúciónak és a genetikai sodródásnak. A variáció mindkét esetben véletlenszerű események eredménye: vagy újonnan fellépő mutációra, vagy új allélt hordozó, bevándorló egyedekre vezethető vissza. A következőkben változások történhetnek az allélfrekvenciákban, ún. allélszubsztitúció (allelic substitution) léphet fel. A genetikai sodródás (drift) és a természetes szelekció közötti különbség próbaköve, hogy az allélszubsztitúció befolyásolja-e a fenotípust, pontosabban a fenotípus fitnesszét. A genetikai sodródásnál nincsen ok-okozati összefüggés az allélvariáció és az egyedek fenotípusos variabilitása (fitnesszvariabilitása) között. A genetikai sodródás ellentétben áll a darwini eszmével, mivel a természetes szelekcióval zajló evolúciónál a folyamat pontosan azért lehetséges, mert az allélszubsztitúció a fenotípus variációjához vezet, és ez a variáció felelős a fitnessz variációjáért. A látványos adaptációk kialakulásában a sodródás szerepe nagy bizonyossággal kizárható.
Vegyünk például egy fűpopulációt, amely olyan környezetben él, ahol magas a herbicidkoncentráció. Ha egy növény hordoz egy allélt, amely herbicidrezisztenciát eredményez, annak az egyednek magasabb lesz a fitnessze, azaz nagyobb valószínűséggel él túl és szaporodik, mint az alléllal nem rendelkező egyedek. A rezisztenciáért felelős allél gyakoribbá válik a következő generációkban, gyakorisága talán elérheti a 100%-ot, azaz fixálódik. Mivel ez a változás nem eredményezett új fajt, az allél migrációval továbbterjedhet a faj további populációiba.
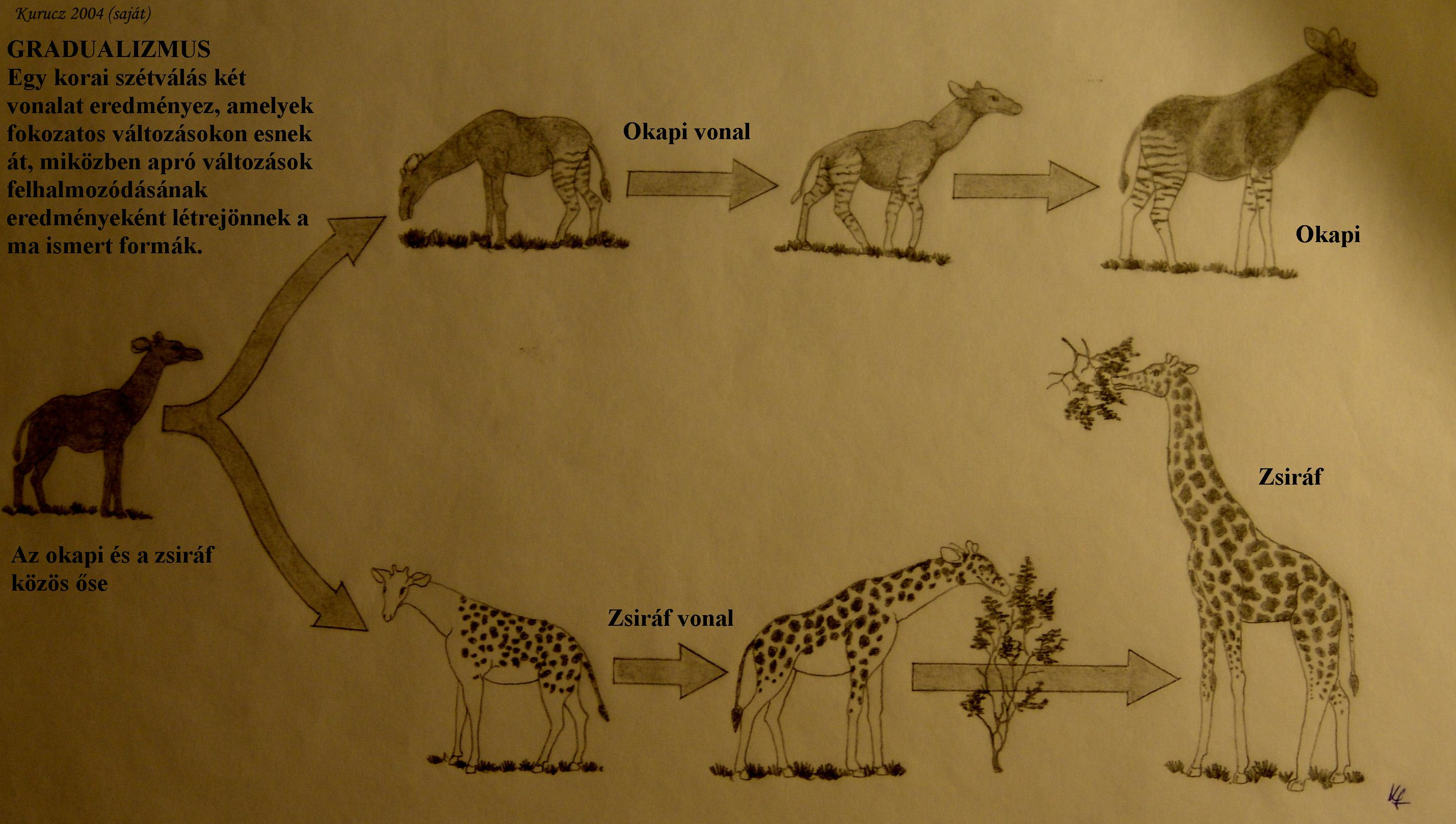
Ez az adaptív forgatókönyv megfelel az anagenezisnek : egy faj adott irányba változik az időben. A kifejezés az ógörögből származik, jelentése: „megint ugyanaz az ág”. Az anagenezis lehet graduális, azaz fokozatos (graduális) és megszakított (abrupt). Ha az új populáció eléggé különböző az ősitől, új nevet lehet adni, az új taxon „kronospeciest” alkot. A kronospeciesekhez vezető anagenetikus folyamatokat gyakorta nevezik filetikus fajképződésnek (phyletic speciation) vagy filetikus evolúciónak.
A filetikus fajképződés különbözik a kladogenezistől, a tulajdonképpeni fajképződéstől (azonos a speciációval), amelynek során egy faj kétfelé ágazik ((3. ábra) . A kladogenezis szintén ógörög eredetű szó, jelentése: „az ág eredete”. Az anagenezishez hasonlóan a kladogenezis is lehet megszakított és fokozatos. A kladogenezis történhet iteratív módon, ilyenkor kládok egy hierarchiája alakul ki.
4.3. ábra - Kladogenezis és anagenezis: az okapi és zsiráf közös ősének populációja kettéválik és differenciálódik. Ez a lépés kladogenezis. A szeparált vonalakon belüli változásokat anagenezisnek hívjuk.
 |
Kritikus lépés a fajképződésben: a génpoolok szeparálódása – a kiindulási faj génpoolja kettéválik – a két független pool allélgyakoriságai aztán az evolúciós ágensek hatására eltérővé válnak – ha elég változás gyűlik fel, új találkozáskor nem képesek géneket cserélni!
Vita tárgyát képezte, hogy a mikro- és makroevolúció folyamatosan átmegy-e egymásba, vagy diszkontinuitás figyelhető meg közöttük. A szintézis során Simpson és Rensch kimutatja, hogy a mikro- és makroevolúciós elméletek összhangba hozhatók. Ehhez azt kell bizonyítani, hogy a diszkontinuitást a fejlődő populációk folyamatos sorozatának végpontjain megfigyelhető különbségek okozzák. Minden makroevolúciós folyamat a populációkban és az egyedek genotípusában játszódik le, ezért mikroevolúció is egyben. A fokozatosság szintén általános jelenség (pl. penicillin-rezisztencia, háziasítás, földrajzi fajképződés), a leletanyagban megfigyelhető diszkontinuitás a véletlen terméke (nem minden köztes állapotról rendelkezünk leletekkel).
Problémát jelent eldönteni, hogy a fajok hasadás vagy leágazás útján jönnek létre.
Dichopatrikus fajképződésnél előfordul hasadás, de a peripatrikus eset (új adaptív zóna, gyors változás, ennek következtében tehát a leágazás is) gyakoribb.
További probléma, hogy az evolúció csak idődimenzióban zajló, lineáris folyamat-e? Ennek megoldására jött létre a szaltációs elmélet (lásd arátákról szóló fejezet). A probléma a diszkontinuitás két értelmezéséből fakad. Elkülönítendő a fenetikus és a taxikus diszkontinuitás. Fenetikus: ugyanazon dém (a helyi populációk összessége) tagjai közötti diszkrét eltérés (pl. egy madár dém tagjainak 12 vagy 10 faroktolla van) (4. ábra). Ha ugyanez az eltérés taxonokat különböztet meg, akkor taxikus diszkontinuitás. A kettő nem egy lépésben megy át egymásba, hanem a szelekció útján (is) ható fajképző evolúció során.
4.4. ábra - Madárfarkak különböző számú tollakkal: a taxikus diszkontinuitás példája. A kolibriknek mindig 10, az énekes madaraknak 12 farktolla van.
 |
A taxonoknak tér- és időbeli korlátai vannak, „minden oldalról szakadékok veszik körbe” őket. Ugyanakkor szülő- és utódfajaikkal kontinuitásban vannak. Új alapító populáció létrejöttekor genetikai átstrukturálódás megy végbe (beltenyésztés, új episztatikus kölcsönhatások megjelenése). Nagy populációk nagy inerciája (tehetetlensége) miatt ott mindez nehezebben történik meg, míg a kis populáció ezt a tehetetlenséget könnyebben megtöri, ráadásul fokozott szelekciós nyomás alatt áll. A leginkább perifériális populációk térnek el legjobban az átlagtól, lásd Melospiza (később) vagy Drosophilák, amelyeknek több száz faja él Hawaiion. A legtöbb perifériális izolátum eltűnik vagy visszaolvad a szülőpopulációba, de ha egy fajban „aberráns” vagyis a többi populációtól látványosan eltérő csoportot látunk, az szinte mindig egy elszigetelt csoport. Üvegnyak-evolúció zajlik: a hirtelen lecsökkenő létszámú populációkban a változatosság egy része elveszik. A hasonlat onnan jön, hogy a populáció létszámát az idő függvényében ábrázolva üvegnyakhoz hasonló képet kapunk. Az elszigetelt csoportoknak is szembe kell azonban kerülniük a nagy, változatos fajokkal, általában nem is tudnak azok elterjedési területeire betörni. A körülmények változásával aztán ez mégis bekövetkezhet, pl. posztglaciális expanziók során.
A makroevolúciós kutatások egyik fő célja, hogy árnyalt képet adjanak az állatvilág leszármazási viszonyairól, elsődlegesen a főbb csoportok (törzsek) kapcsolatairól. Ezen a területen hatalmas előrelépés történt a kétezres években, az élet fája, legalábbis főbb ágait tekintve, köszönhetően elsősorban a genomikai és bioinformatikai kutatások robbanásszerű fejlődésének, kialakult. Az élet egyik fő mintázata tehát kirajzolódott, mint azt a későbbiekben majd felvázoljuk.
Ritkán gondolunk rá, de a több, kisebb klád előfordulása a nagyobb kládokon belül is a makroevolúciós mintázatok közé tartozik. Például minden hajtásos növénynek közös sajátságai vannak (pl. a hím ivarsejteket hordozó pollenszemek, a megtermékenyítés után a női reproduktív szervekben maradó női gaméták, a nőivarú növénytől származó szövetburokba csomagolt embrió, stb).
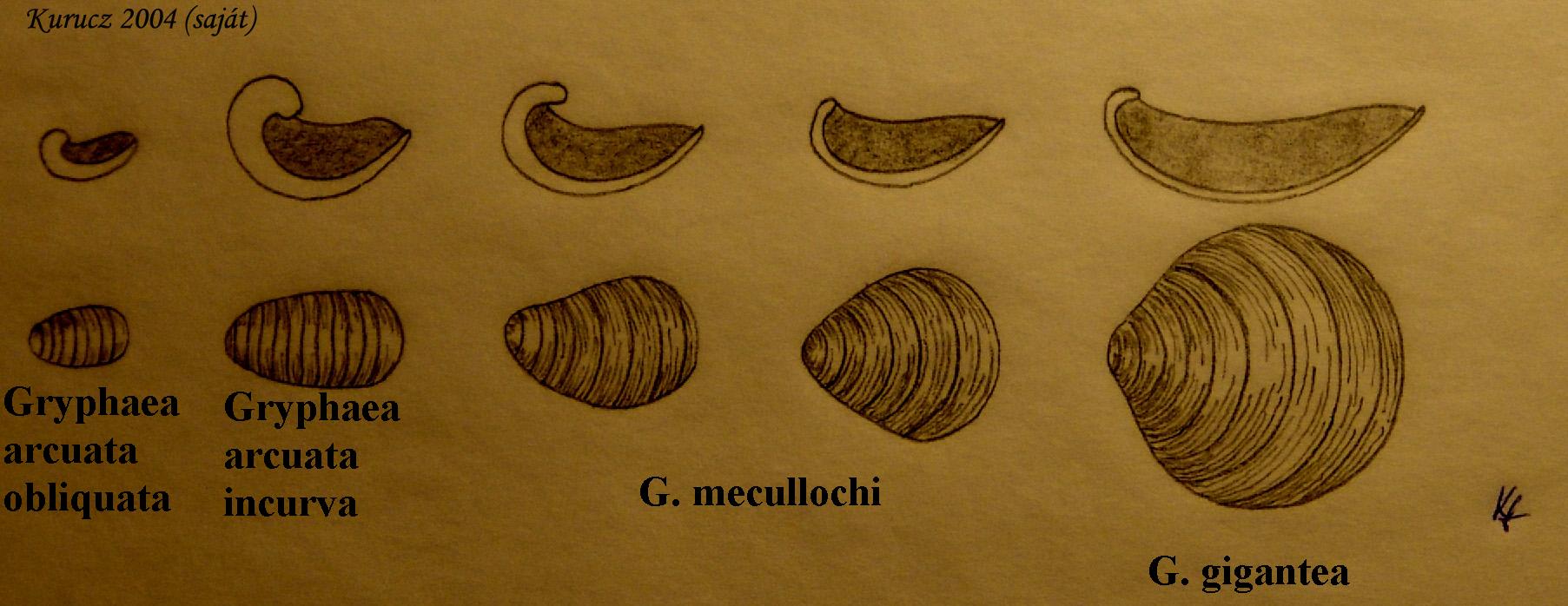
A hajtásos növények óriási kládja felosztható kisebb kládokra, mint az ősi nyitvatermők és az „újabban” megjelenő zárvatermők, amelyek 200-150 millió évvel ezelőtt jelentek meg, és a nyitvatermőktől sok vonatkozásban különböznek azon felül is, hogy virágaik vannak. Specializált vízszállító sejtjeik vannak, és specializált endospermium szövet látja el az embriót a magon belül. A zárvatermők ökológiai szempontból is drámai eltéréseket mutatnak: míg a nyitvatermők pollenjét és magvait a szél terjeszti, a zárvatermők jobban szorítkoznak más tényezőkre (rovarok, más állatok) ezeknél a funkcióknál. A zárvatermők nem teljes mértékben szorították ki a nyitvatermőket, de dominánsakká váltak bolygónk sok területén. Ahogy terjedtek, a zárvatermők hatalmas mértékben diverzifikálódtak: új fajok és magasabb rendszertani egységek jelentek meg. Az adaptív radiáció számos jelleg komoly mérvű megváltozásához vezetett: makroevolúciós változás volt.
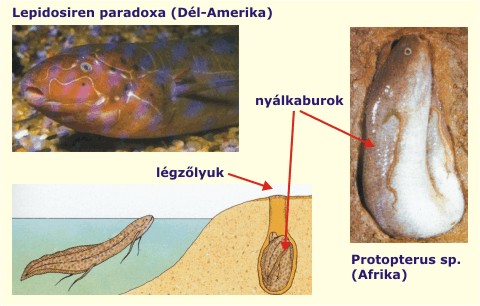
Egy másik drámaian látványos, makroevolúciós jelenség az eleven fosszíliák léte: ezek olyan modern fajok, amelyek csak keveset változtak a távoli evolúciós múltban élő őseikhez viszonyítva. A morfológiai szempontból statikusnak tekinthető élőlények együtt élnek a később kifejlődött élőlényekkel. Klasszikus példának számít például az atlanti tőrfarkú, tüdőshalak (5. ábra), pajzsosrákok, Nautilusok (6. ábra) és a szitakötők. Az élő fosszíliák sokszor olyan nagyobb rendszertani egységek tagjai, amelyek kivételükkel teljesen kihaltak. Az eleven fosszíliák léte a csaknem nullára lassult rátájú evolúciós változás, a sztázis drámai megnyilvánulása.

Fontos makroevolúciós kérdés: miért diverzifikálódik némelyik klád látványosan az idők folyamán, míg mások változatlanul maradnak, megint mások kihalnak. A kutatók egy része úgy gondolja, hogy fajszelekciós folyamatok (species sorting és species selection névvel tesznek finom különbségeket, mint látni fogjuk) játszhatnak közre. A fajszelekció koncepciója arra épül, hogy a faj születése (speciáció) és kihalása analóg az egyed születésével és halálával. Ahogy az egyed fitnessze függ az adaptációitól, annak a valószínűsége, hogy a faj túlél, vagy kihal, bizonyos sajátságoktól függ, amelyek a fajnak, mint összességnek a sajátságai. Ezek a sajátságok különböznek az egyes egyedek sajátságaitól, mint pl. a hatékony tovaterjedési képesség, ehelyett a faj globális sajátságain múlnak, mint a széles földrajzi elterjedés. A fajok kiválogatódása (az angol fogalom, amit ilyen széles értelemben használnak: species sorting) nyilvánvalóan lehetséges, ha a fajok között ilyen természetű különbségek vannak. A fajszelekció (species selection) az előzőnél specifikusabb mechanizmus, akkor működik, ha a fajokra jellemző globális sajátságok átadódnak az új fajnak a speciáció során: ha pl. az új faj szükségszerűen nagy elterjedésű lesz, egyszerűen azért, mert a faj, amelyből származik, szintén széles elterjedésű volt.
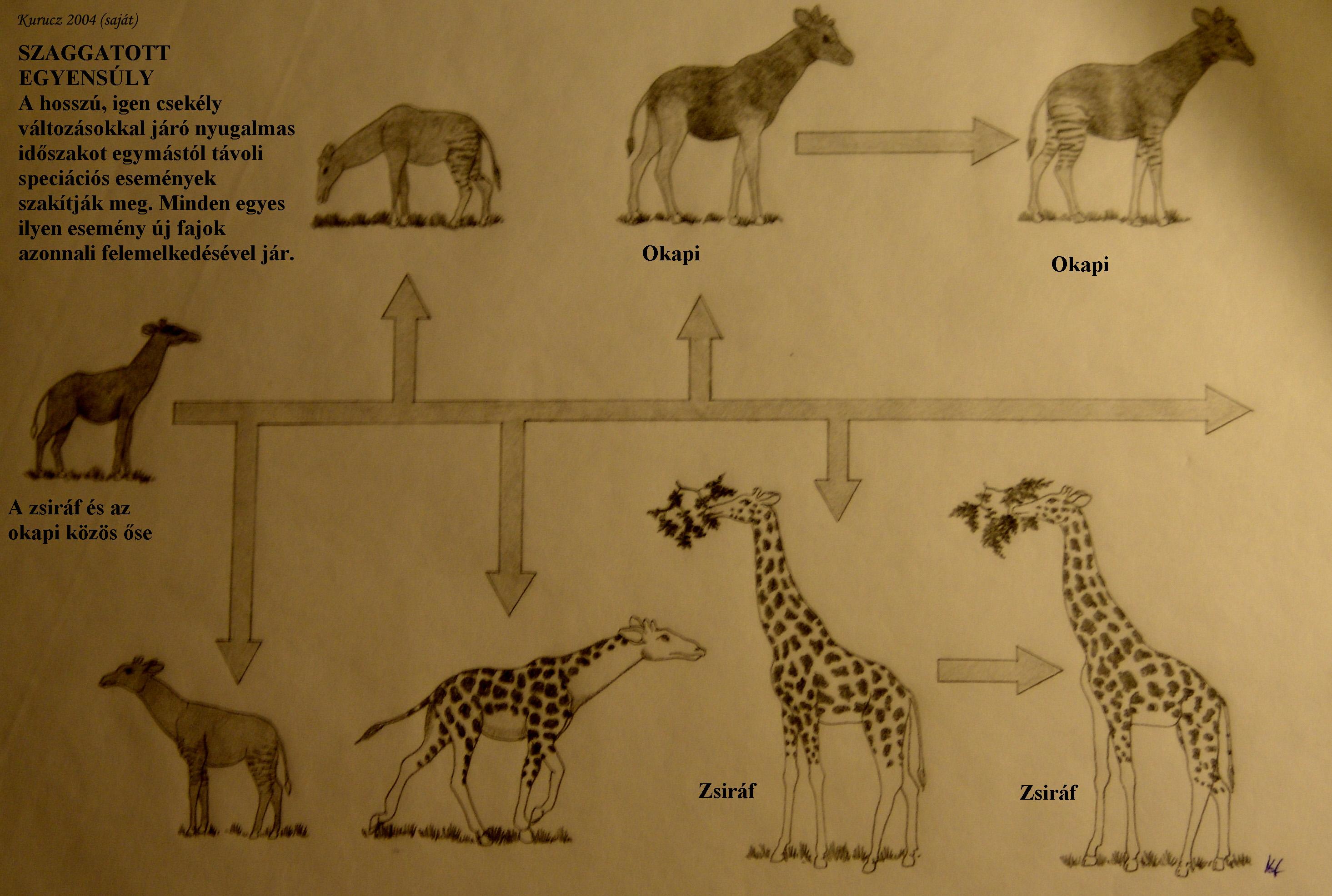
A makroevolúció egyik legfontosabb kérdése a szaggatott egyensúlyé (punctuated equilibrium). Eldredge és Gould 1972-ben vezette be az elméletet, két megfigyelésre építve:
(1) A legtöbb fosszilis faj a látszat szerint átmenet nélkül bukkan fel, és azután együttesen fordul elő „elődeivel” azaz a fajjal, amelyből származik.
(2) A kladogeneziskor megfigyelhető változások gyorsak és kivételesek, a fajok egyébként legtöbbször évmilliókig változatlannak tűnnek, azaz sztázisban vannak.
E két, egymással szorosan összefüggő megfigyelés kétségessé tette a Darwin által is favorizált filetikus gradualizmus eszméjét, azonban szépen összeilleszthető az Ernst Mayr által javasolt perifériás izolációs modell néven ismert fajképződési mechanizmussal. A modell lényege: a speciációban gyakran játszik szerepet egymástól földrajzilag elkülönült populációk divergenciája (7. ábra). A speciáció után a két faj (a „régi” és az újonnan keletkezett) akkor is egymás mellett létezhet, ha a geográfiai elkülönültség megszűnik. A modell szerint lezajló speciációs események csak elenyésző valószínűséggel maradnak fenn a fosszilis anyagban, különös tekintettel a folyamat helyi korlátozottságára.
A szaggatott egyensúly modell (8. ábra) gyökeresen különbözik a filetikus gradualizmustól , és új utakat nyitott az evolúció kutatásában. Alapvető kérdéssé vált a fajok szétválásának mikéntje, és hogy mire vezethető vissza a sztázis. A stabilizáló szelekció akadályozná meg a faj morfológiájának gyors változásait az idők folyamán? Vagy vannak olyan genetikai és fejlődési korlátozó tényezők („constraints” az angol nyelvű szakirodalomban), amelyek egy adott irányba terelik, vagyis „kanalizálják” az evolúciós változást?
Mikroevolúciós változások megfigyelhetők mesterséges körülmények között: akár a laboratóriumban, akár a mezőgazdaság keretein belül. A mikroevolúciós ráták így közvetlenül és pontosan számszerűsíthetők, és összevethetők a fosszilis anyagból kikövetkeztetett evolúciós rátákkal. A paleontológia módszertárának fejlődése szigorú teszteket tesz lehetővé, és a ráták becslése sokat változott az utóbbi időben, de a mikro-, és makroevolúciós ráták összevetésére való törekvés nem új. Simpson karrierje jelentős részét a makroevolúciós ráták a fosszilisanyag alapján történő kvantifikálásának szentelte. Ő volt a darwin, mint evolúciós egység bevezetésének úttörője. Maga az egység a kiváló elméleti genetikus J. B. S. Haldane (9. ábra) által javasolt kvantitatív módszeren alapult. Haldane módszere tette lehetővé, hogy egy adott jelleg evolúciós rátája összevethető, még akkor is, ha a jelleg mérete drámaian különbözik a rokon élőlényekben: összevethető pl. a kolibri és a strucc combcsontjának evolúciós rátája. A módszer megengedi azt is, hogy egy adott vonal evolúciós történetének különböző időpontjaiban hasonlítsák össze az evolúciós rátákat: Simpson példájában a modern lófélék fogainak a sokkal kisebb, csakis fosszilis anyagból ismert kisebb lovakéval való összevetése is lehetséges. (Mint látni fogjuk az evolúciós ráták részletesebb elemzésekor, az újabb egység, a haldane már elfogadottabb.)

A módszernek megvannak a maga korlátai: A jelleg annyit változhat a hosszú távú evolúció során, hogy teljesen transzformálódik. A fog agyarrá alakulhat, vagy méretben annyira drasztikusan lecsökkenhet, hogy eltűnik. A módszer mégis gyakorta sikeres, és egy, a nyolcvanas években végzett, kiterjedt összehasonlító vizsgálat szerint a „kísérletes” evolúciós ráták 10 000- 100 000-szer nagyobbak, mint a fosszilis anyag alapján számított evolúciós ráták. Nyilvánvaló, hogy a mikroevolúciós mechanizmusok elégségesek a makroevolúciós változások magyarázására.
Gyakori kritika volt ezzel a megközelítéssel kapcsolatban, hogy a laboratóriumi környezetben a szelekciós nyomások szélsőségesen mesterségesek a természetben uralkodó szelekciós viszonyokhoz képest. A fenti állítás cáfolására az 1980-90-es években kiterjedt vizsgálatokat folytattak annak megállapítására, milyen gyorsan hat a természetes szelekció természetes körülmények között. Alapos és viszonylag hosszútávú vizsgálatok folytak a trinidadi guppikkal (Poecilia reticulata), amelyek meggyőzően bizonyították, hogy gyors mikroevolúciós változások lehetségesek a természetben is 10. ábra.

A trinidadi guppik kétféle, elkülönülő folyószakaszban élnek: (1) ragadozóktól csaknem mentes és (2) jelentősen magasabb predációs rátájú szakaszokon.
Mivel a vízesések és a sellős folyószakaszok gyakorta korlátozzák mind a guppik, mind ragadozóik mozgását, a két féle guppi populáció izolált egymástól. Ha a kétféle típusú populációból származó egyedeket párhuzamosan nevelnek fel laboratóriumi körülmények között, a magas predációjú környezetből származó egyedek korábban, kisebb mérettel érnek, ami a nagyobb példányokat előnyben részesítő predátorok elleni védekezés. A kutatók feltételezték, hogy szelektálni tudnak olyan halakra, amelyek később és nagyobb mérettel érnek, ha áthelyezik őket a folyó alacsony predációjú szakaszaiba. A 11 évet felölelő, a populációk áthelyezésén alapuló vizsgálatok során az áthelyezett populációkat hasonlították össze az eredeti, „kontroll” populációkkal, és azt találták, hogy az áthelyezett populációk pontosan a megjósolt irányba fejlődtek: később, és jelentősen nagyobb mérettel érték az érettséget.
A guppikkal végzett kísérletek markánsan megmutatták, evolúciós mértékkel milyen „képtelenül” gyorsan reagáltak az áthelyezett populációk az új szelekciós körülményekre, adott esetben a gyengébb predációra. Az evolúciós változás sebessége ugyanolyan gyors volt, mint azt a korábbi laboratóriumi kísérletek során tapasztalták, „mesterséges” szelekciós körülmények alkalmazásakor. A következetes mintázat és a gyors változási ráta elégséges azoknak a mintázatoknak a magyarázására is, amelyeket a sokkal hosszabb időintervallumot felölelő fosszílis anyagnál megfigyelhetők. A guppi kísérletek alapján úgy becsülték, hogy a tanulmányaikban tapasztalt ráta 10 000- 1 000 000-szor nagyobb, mint a fosszilis anyag alapján számították.
A kísérletek tehát meggyőzően bizonyították, hogy a természetes szelekció gyors evolúciós változásokat okozhat. Miután nincsen ismert evolúciós mechanizmus, amely felelős lehet gyors evolúciós változásért, ésszerű következtetés, hogy a mikroevolúciós változásokkal magyarázhatók a fosszíliák evolúciós változásai.
A fenti tanulmányok nem zárják ki azt a lehetőséget, hogy új makroevolúciós mechanizmusokat fedeznek majd fel.
Két, további megjegyzés kívánkozik még ide: (1) a kísérletek nem vezettek kladogenezishez, így nem mondanak sokat a speciációról és a makroevolúcióról.
(2) Miért zajlik annyira lassan a fosszilis anyagot alapul véve az evolúció, ha potenciálisan sokkal gyorsabban is folyhatna? A kutatók szerint a környezeti tényezők mindenféle következetesség nélkül változnak, ami olyan szelekciós körülményekhez és a rájuk adott olyan válaszokhoz vezet, amelyek szintén változnak az időben, de végeredményben nincsen mérhető változás. Bár a felvetés legalábbis plauzibilis, a kutatók egy része úgy gondolja, hogy a mikroevolúciós mintázatok nem elégségesek minden makroevolúciós változás maradéktalan magyarázatára.
A makro-, és mikroevolúció mechanizmusai körüli viták nyugvópontra jutásához mindenképpen szükséges a perifériás izolációs modell további részletes elemzése. A filetikus gradualizmus elméletével összhangban az izolált populációkban alakulnak ki legnagyobb eséllyel a genitáliák és a párosodási viselkedés változásai, nagy valószínűséggel reproduktív izolációt és speciációt vonva maguk után. További fontos körülmény, hogy az izolált populációk gyakorta vannak kitéve újfajta, stresszel járó szelekciós körülményeknek. Végezetül, ha az izolált populációk kicsik, a genetikai sodródás (ami random változásokat jelent az allélfrekvenciákban), fontos mechanizmusává válhat a nem adaptív, mindazonáltal mikroevolúciós változásoknak.
A makroevolúcióval foglalkozó tudósok közül többen is felvetették, hogy a perifériás populációkban még különösebb jelenségek is lejátszódnak. Az új, szokatlan környezet nem csak új szelekciós tényezőket jelent, hanem a normális, homeosztázist biztosító, a stabil fenotípust fenntartó mechanizmusok is felborulnak. Továbbá, miután a genetikai sodródás működhet az ilyen populációkban, akár nagyhatású és káros mutáció maradhat fenn, sőt, terjedhet el, akár széleskörűen. Ha a végeredmény nem kihalás, új faj jelenhet meg, drámaian különböző, és nem is feltétlenül adaptív jellegekkel. Ilyen szaltációs (ugrásszerű) vagy makromutációs változásokat tételeztek fel, vizsgáltak, és vetettek el a huszadik század során, annak első felében, de a huszonegyedik század elején újabb eredmények és módszerek arra sarkallták a kutatókat, hogy a régi eszméket legalábbis átgondolják.
Az 1950-es években a Drosophila melanogasteren végzett kutatások feltárták, hogy (i) a mutáció, (ii) vad típusú egyedek egyszerű éteres sokkja, vagy hősokkja a felnőtt életet megelőző életszakaszokbana a felnőtt egyed morfológiájának, például a szárny erezetének megváltozását okozhatja, sőt akár homeotikus fenokópiákat is eredményezhet. Mesterséges szelekciós kísérlet során olyan populációt állítottak elő, amely tendenciózusan mutatott ilyen, a környezet iránt indukált ún. fenokópiákat, azaz a mutánsnak megfelelő fenotípusokat vad genotípusú legyeken. A mesterséges szelekcióval emelni lehetett a fenokópiák gyakoriságát, sőt, olyan egyedek is létrejöttek, amelyek az abnormális fenotípust mutatták hősokk nélkül is.
Eltelt 40 év, és a kilencvenes években a fenokópiákkal végzett kísérleteket az University of Chicago molekuláris biológusai ismételték meg. Számukra már több ismeret állt rendelkezésre: tudták, hogy ha hősokkot alkalmaznak egy ún. hősokk fehérjének, a Hsp90-nek szerepe van a homeotikus állapot fenntartásában, vagyis a káros hatások „pufferelésében.. Ha a fehérjét akár egy mutáció, akár gátló vegyülettel funkcióképtelenné tették, morfológiai változások - köztük a szárnyi vénák mintázatának megváltozása - jelentek meg a Hsp90 funkciót nélkülöző legyekben, míg a vad típusú legyek a hősokk mellett is megmaradtak vad típusúaknak. Akárcsak a régebbi tanulmányok során, mesterséges szelekcióval elérték, hogy a fenokópiák gyakorisága nőjön, és végül kifejeződjenek akkor is, ha a Hsp90 funkcióképes fehérje jelen van az egyedekben.
Összefoglalva, az eredmények arra utalnak, hogy a genetikai variáció „rejtett” forrása van jelen a genomban, és az új, és/vagy stresszes környezet feltárja ezt a variációt. A megfigyelés kiemelt jelentőséggel bír, ugyanis a variáció a genetikai és evolúciós változás nyersanyaga. Érdekes továbbá, hogy az eredmény szépen egybevág a szaggatott egyensúly elmélet szélsőségesebb felvetéseivel, mint azt már fejtegettük, és a makromutációkra alapozott evolúcióval is viszonylag könnyen kapcsolatba hozható (ld. a későbbieket).
Egy másik, ugrásszerű evolúciós változásra ad példát a kukorica (Zea mays) két variánsa: a mindenki által ismert domesztikált kukorica, és legközelebbi vad rokona, őse, a teozinte.
A két variáció, bár kereszteződnek, külső morfológiájukat tekintve drasztikusan eltérnek. Éréskor a teozinte magvai kisebb, megnyúlt, könnyen széteső és magjait szétszóró „csövet” alkotnak. Ezzel szemben a kukorica csövei hosszabbak, vaskosabbak, és nehezebben esnek szét. A teozinte és kukorica különbözik a teljes növény megjelenésében is. A kukorica magas növény, amelynek szára nem ágazik el. A hím virágzat, a címer a növény csúcsán helyezkedik el, míg a női virágzatok (a csövek) a szártengely mentén, az egyes levelek és a szár által körülvett térben.A teozinténél a hosszú elágazások ugyanott nőnek, mint a női virágzatok, egy bokrosabb növényt hozva létre. A bokros teozinte növény reproduktív szempontból sikeresebb, mert minden oldalágon további női virágzatok jöhetnek létre. Érdekes, hogy a teozinte növények csak akkor nőnek a leírt módon, ha egy napos, nyílt terepen kelnek ki. Ha nagyobb sűrűséggel vetik őket, hasonlóan a domesztikált kukoricához, a teozinte hosszabbra növeszti fő szárát, és hasonlóbb lesz a kukoricához. A kukorica akkor sem nő bokrosan, ha magában növesztik (11. ábra).

A kukorica evolúciójának kutatóit régóta foglalkoztatta a teozinte és kukorica közötti, lényeges különbség genetikai alapja, és hogy a változások vajon milyen gyorsan jelentek meg. Az egykoron szélsőségesnek tekintett gondolat, hogy mindezen változások egyetlen „makromutációnak” köszönhetők, már jó egy évtizede meggyőző igazolást nyert (Lásd 1. fejezetben is, a kukorica háziasításával kapcsolatban). A teozinte branched 1 (tb1) génnek két allélja van, a „teozinte” allél és a „maize” (=kukorica) allél. A teozinte és a kukorica keresztezhető, így a kutatók keresztezésekkel előállítottak teozinte növényeket „maiz” alléllal, és kukorica növényeket „teozinte” alléllal. Az ilyen növények fenotípusa megmutatta, hogy a tb1 gén alléljai felelősek a legtöbb morfológiai különbségért.
A tb1 gént azonosították és szekvenciáját meghatározták. Úgy tűnik, a gének kódoló szakaszában nincsenek a funkció szempontjából lényegi különbségek a vad típusú és a domesztikált formára jellemző allél között, de a génhez kapcsolódó („flanking”) régiókban jelentős szekvenciakülönbségeket találtak. Ezek a régiók cisz-regulátor régiók, amelyek szabályozzák hol, mikor, és hogyan fejeződjön ki a gén, amelynek terméke jelentős hatással van a növény fejlődésére. Az is valószínű, hogy az új, ez esetben mezőgazdasági környezet segítette a domesztikált kukorica túlélését akkor is, ha a tb1 mutáns allél a teozinte átmeneti, adaptív fenotípusát jelenti .
A homeotikus mutációk sokszor „szörnyetegeket” hoznak létre (12. ábra), amelyek kulcsjelentőségűek voltak a toolkit feltárásában, és semmiképpen sem tévedünk nagyot, ha őket „makromutációként” kezeljük, mert látványos, sokszor lényegi változásokat okoznak az állatok testtervében. Miután kulcsjelentőségűek, számtalan helyen foglalkozunk velük a következő fejezetekben.

A kutatók egyre többet tudnak meg a gyors makroevolúciós változásokról. Fontos tudatában lennünk, hogy a makroevolúciós mechanizmusok megismerése nem teszi kérdésessé alapos ismereteinket a fokozatosságon alapuló mikroevolúciós mechanizmusokról. Általánosan elfogadhatjuk, hogy a makroevolúciós és mikroevolúciós változások mechanizmusok nem egymást kizáró magyarázatok az evolúciós változásra, hanem hierarchikus kapcsolatban állnak, és együtt erősebb és mélyebb magyarázatot adnak a biológiai diverzitás evolúciójára.